

CAPÍTULO 7
VEGETACION NATURAL DE LA ZONA DE IQUITOS
Kalle Ruokolainen y Hanna Tuomisto
Se estudió la composición florística del bosque en base a tres grupos de plantas: (1) árboles (>2,5 cm DAP), (2) Melastomatáceas (una familia mayormente de arbustos), y (3) pteridófitas (helechos y plantas afines). Los árboles fueron inventariados en 11 sitios diferentes, dentro de los cuales se reconocieron un total de 15 sectores de muestreo basándose en características del terreno y de los suelos. En cada sector, se levantaron 2-4 parcelas (20 x 20 m 6 25 x 25 m). Melastomatáceas y pteridófitas fueron inventariadas en los mismos 15 sectores que los árboles, y en 17 sectores adicionales en sitios diferentes. Las Melastomatáceas y pteridófitas se muestrearon en las mismas parcelas donde se hizo el inventario de árboles, y además se muestrearon a lo largo de transectos (2 x 500 m, 5 x 500 m 6 5 x 1300 m por sitio). En el inventario se encontraron un total de 1416 especies de árboles, 134 especies de Melastomatáceas y 152 especies de pteridófitas. Se recolectaron muestras de suelo para analizar sus características físicas y químicas. También se utilizó una imagen de satélite Landsat TM para obtener información sobre la reflectancia de luz en cada uno de los sectores de estudio. Este solo se pudo hacer en 23 de los 32 sectores porque los demás estaban cubiertos por nubes o presentaban características obvias de intervención humana que alteraba las reflectancias. Las semejanzas florísticas entre los sectores de estudio se calcularon como porcentaje de especies compartidas entre dos sectores. Eso se hizo separadamente con árboles, Melastomatáceas y pteridófitas. Con las características de suelo y de imagen de satélite, las diferencias se calcularon mediante la aplicación de las distancias euclidianas.
Los resultados de los análisis indican que los tres grupos de plantas presentan patrones muy parecidos de similitud entre los sectores de muestreo. Los patrones florísticos también están correlacionados con los patrones obtenidos con características de los suelos, especialmente con la textura y los cationes Ca, K y Mg. La información de las reflectancias de luz también produce un patrón de semejanzas y diferencias entre los sectores que es parecido a los patrones florísticos y edáficos. Estos resultados sugieren que es posible utilizar las Melastomatáceas y pteridófitas como indicadores de patrones florísticos generales en los bosques amazónicos, y también para indicar aproximadamente algunas características del suelo. Además, parece posible identificar diferentes tipos del bosque en base a la información registrada en las imágenes de satélite.
En: Kalliola, R. & Flores Paitán, S, (eds.) 1998. Geoecología y desarrollo Amazónico: estudio integrado en la zona de Iquitos, Perú. Annales Universitatis Turkuensis Ser A II114:253-365.
El clima en la zona de Iquitos es siempre húmedo sin mucha variación de temperatura o precipitación entre los diferentes meses del año (véase Capítulo 3). En términos generales se describe (Holdridge 1967, Walter & Breck1e 1984) que este tipo de clima soporta una vegetación exuberante de bosque alto, cuyo dosel llega hasta la altura de aproximadamente 30-40 m o más, y a intervalos existen además unos árboles gigantescos tanto en diámetro como en altura. Los árboles están cargados con epífitas y lianas, y todo el verdor está compuesto por una cantidad muy alta de diferentes especies de plantas. Esta es la percepción estereotípica de los bosques húmedos tropicales de elevaciones bajas.
Evidentemente esta visión general tiene cierta razón en consideraciones de gran escala, pero es igualmente claro que en una escala más detallada se puede diferenciar una gran variación tanto en la estructura como en la composición florística de la vegetación. En Amazonía, una de las divisiones más drásticas del ambiente está constituida por la inundación de los ríos, y la distinción entre tierra firme y áreas inundadas se ha incluido ya en las primeras descripciones escritas de la Amazonía.
Las áreas inundadas están fuera del enfoque de este articulo, y para aquellos quienes quieren introducirse a la vegetación inundable en la Amazonía o en la Selva peruana hay varias otras obras de referencia (por ejemplo: Prance 1979, Encarnación 1985, Kubitzki 1989, Klinge et al. 1990, Parodi & Freitas 1990, Kalliola et al. 1991, Kvist et al. 1995).
En áreas no inundables (tierra firme) en la Amazonía, la vegetación varía principalmente según las condiciones climáticas y edáficas (Pires & Prance 1985, Prance 1989, Duivenvoorden & Lips 1993, ter Steege et al. 1993, Ruokolainen et al. 1997). En la zona de Iquitos la variación geográfica en el clima es, al menos hasta donde se conoce, muy exigua y por eso las características del suelo tienen la mayor importancia en determinar la vegetación. La distinción más clara en la tierra firme se hace entre vegetación sobre arena blanca y vegetación sobre suelos limoso-arcillosos.
En la zona de Iquitos se han podido distinguir tres tipos diferentes de vegetación en arena blanca: varillal seco, varillal húmedo y chamizal. El varillal seco contiene un bosque más alto y con más especies que el varillal húmedo, el cual se caracteriza por una capa gruesa (hasta 50 cm) de raíces y material orgánico (Ruokolainen & Tuomisto 1993, Tuomisto & Ruokolainen 1994). En el chamizal el dosel de la vegetación se encuentra a tres metros o menos y el aspecto es de matorral (Encarnación 1985). En casos extremos, la altura de la vegetación no sobrepasa un metro y crecen líquenes en el suelo (Ruokolainen & Tuomisto 1993). Los nombres varillal y chamizal. son los nombres que los campesinos locales utilizan para los tipos de vegetación en arena blanca, aunque posiblemente en sentidos un poco diferentes. Por lo menos la gente que vive a lo largo de la carretera Iquitos-Nauta aplica el nombre chamizal a bosque de varillal húmedo con una capa muy gruesa y floja de raíces (S. Flores Paitán, comunicación personal).
Es probable, aunque falta todavía la verificación, que los tres tipos de vegetación en arena blanca están determinados por la variación en el drenaje controlado por una capa impermeable del suelo (que puede estar conformada por arcilla dura o es el horizonte espódico con alta concentración de hierro o manganeso, véase Capítulo 5). En varillal seco aparentemente no existe una capa impermeable, o si existe se sitúa a una profundidad que permite un buen drenaje, todo el tiempo, para las raíces de las plantas.
En el varillal húmedo la capa impermeable está tan cerca de la superficie que después de mucha lluvia el suelo queda saturado por agua. También en el chamizal las lluvias producen saturación de agua en el suelo, o hasta inundación. Por otra parte, después de algunos días de sequía el suelo puede secarse completamente porque hay muy poca tierra sobre la capa impermeable,.y por eso una baja capacidad de retención del agua. Aparentemente los cambios drásticos entre inundación y sequía completa hacen que las condiciones del chamizal sean imposibles para el crecimiento de árboles, y solo plantas de menor tamaño pueden soportarlas.
Encarnación (1985) ha identificado seis tipos diferentes de vegetación de tierra firme en suelos que no son de arena blanca. El tipo más especial es sin duda supaichacra que está formado casi solo por dos especies arbóreas mirmecófilas, Duroia hirsuta y Cordia nodosa. En partes mal drenadas se encuentran los aguajales de altura, los cuales son caracterizados por la abundancia de dos palmeras grandes, Mauritia flexuosa (aguaje) y Oenocarpus bataua (ungurahui). Yarinal de altura se llaman a las manchas de bosque que tienen drenaje lento y presencia de Phytelephas macrocarpa (yarina).
Los terrenos bien drenados presentan, según Encarnación (1985), tres tipos de vegetación: bosque de terraza, bosque de colina y bosque de quebrada. ' Esta división es principalmente topográfica, como los nombres mismos lo revelan, y falta documentar posibles diferencias en la estructura y composición florística.
Además de dichos seis tipos de vegetación, existen en valles mal drenados, dentro de áreas relativamente pequeñas, pantanos casi abiertos sin árboles pero con hierbas grandes de las familias Araceae y Zingiberaceae (Ruokolainen & Tuomisto 1993).
Los bosques de colina y bosques de terraza cubren juntos más del 90% de la superficie de tierra firme en la zona de Iquitos, y por eso se podría pensar que la vegetación sea relativamente homogénea en grandes extensiones. Sin embargo, hay que reconocer que la idea de homogeneidad en realidad nace de la falta de documentación y conocimiento exacto. Simplemente no han existido observaciones que hubiesen podido ni confirmar ni efectivamente rechazar el concepto de homogeneidad. Solo muy recientemente se ha podido documentar (Gentry & Ortiz 1993, Ruokolainen et al. 1997) que según la fertilidad de los suelos se pueden dividir las áreas de bosque de colina y de terraza en por lo menos dos unidades que tienen composición distinta de especies.
La falta de información sobre la variación de la vegetación es un obstáculo para la conservación y el uso sostenible de este ambiente. Como existe una presión creciente ejercida por la colonización y utilización de madera, caza, frutos, fibras y hasta genes presentados por la flora y fauna de la región, se necesita urgentementela información correspondiente. Evidentemente, uno de los factores que ha obstaculizado la acumulación de datos de campo es la riqueza excepcional de especies. Es posible encontrar hasta 300 especies de árboles en una hectárea (Gentry 1988), la flora de algunas reservas en la zona de Iquitos reconoce casi 3000 especies (Vázquez 1997) y seguramente existen todavía varias especies sin colectar y/o sin nombres científicos. La diversidad de la flora hace que los trabajos de colecta e identificación de las muestras botánicas requieran bastante tiempo y recursos.
En este trabajo hemos tratado de evitar tales problemas de manera tal que se ha concentrado los estudios en dos grupos de plantas (pteridófitas y Melastomatáceas) para que se pueda investigar más sitios en el campo. La representatividad de los resultados en relación con la composición florística de árboles se verificaron en un número más restringido de sitios (Ruokolainen et al. 1997). Una discusión más profunda sobre la idea de utilizar plantas indicadoras de tipos de bosque se presenta en el Capítulo 13.
Otra manera de cubrir un área mayor y extrapolar los resultados del trabajo del campo es la utilización de imágenes de satélite Landsat TM. Las imágenes de satélite consisten de una red de células o pixeles, de 30 m x 30 m, que cubren toda la tierra. Para cada pixel el satélite registra la intensidad de reflectancia de luz solar en siete bandas, cada una posee una longitud de honda específica desde el azul hasta el infrarrojo. La intensidad de la luz reflejada varía según las características físicoquímicas del terreno, tales como la estructura del dosel de la vegetación, la actividad fotosintética, el color y estructura de las hojas, la humedad del suelo y otros factores. Se puede pensar que cada pixel presenta un resumen medioambiental y la tarea del investigador es hallar la significación y la explicación biológica de dichos resúmenes.
La zona de Iquitos es especialmente interesante para tales estudios, porque presenta una variación amplia en condiciones edáficas, y varios tipos de bosque natural ocurren en la zona. La caracterización y el mapeo de estos ecosistemas diferentes también es muy urgente: hay mucha presión para extender el área cultivada, lo que solamente se puede hacer de una manera rentable y sostenible si la planificación del uso de la tierra está basada en el conocimiento de las condiciones ecológicas de la zona. Al mismo tiempo, la conservación de los recursos naturales depende del conocimiento adecuado de la ubicación y características de los bosques diferentes.
PARCELAS DE COMPOSICIÓN FLORÍSTICA DE ÁRBOLES
En la selección de los lugares para el trabajo del campo, se trató de muestrear áreas que presentan características diferentes en las imágenes de satélite, para averiguar si en la realidad también son distintas. También se trató de cubrir el área de estudio tan completamente como fuese posible, pero obviamente había que restringir el muestreo a aquellas áreas que fueran accesibles a través de los ríos o por la carretera.
Se establecieron parcelas en 11 sitios diferentes (Figura 7.1), en tres de los cuales existen tipos diferentes de terreno que fueron muestreados separadamente como sectores independientes de muestreo. Con algunas excepciones, en cada sitio se establecieron cuatro parcelas de 20 m x 20 m cada una. Las parcelas se establecieron a lo largo de un transecto de 500 6 1300 m de largo (ver abajo) de tal manera que dos de las parcelas se localizaron en las laderas de colinas y dos en las quebradas de valles. De esta forma, se trató de cubrir la variación topográfica en cada sitio para conseguir una idea más completa de la composición de especies a una escala regional. En las áreas de arena blanca tal distribución topográfica de las parcelas no fue posible porque la arena blanca solamente se encuentra en las colinas. Además, en algunos sitios en terrazas fluviales recientes (Mishana 1 y Momón l), el terreno era casi completamente plano y consecuentemente las parcelas en esos lugares tuvieron que representar la misma posición topográfica. Al elegir los lugares exactos de las parcelas, se trató de evitar claros recientes.
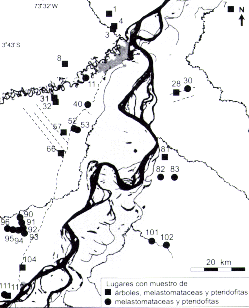
En dos sitios (Mishana y Momón) se establecieron parcelas de tamaño 25 m x 25 m. Las parcelas en Momón se dividieron entre dos tipos de terreno: colinas y planicies, cada uno de los cuales fue muestreado con dos parcelas. En Mishana, se establecieron cuatro parcelas en terraza y cinco en arena blanca o bosque de varillal (Encarnación 1985). Para los otros inventarios, el tamaño de las parcelas fue reducido a 20 m x 20 m para poder muestrear un mayor número de sitios. En 13 de Febrero, el terreno consistió tanto de suelo arcilloso como de suelo arenoso, y cada tipo de suelo fue muestreado con dos parcelas. En los demás sitios, se establecieron cuatro parcelas. Así, tomando en cuenta las divisiones internas de algunos lugares de muestreo, se obtuvieron inicialmente 14 unidades de terreno (11 sitios de muestreo, de los cuales Momón, Mishana y 13 de Febrero incluyeron dos unidades de terreno cada uno).
Posteriormente, en base de las diferencias y semejanzas florísticas entre las parcelas (véase la parte de Resultados abajo), se consideró necesario interpretar las parcelas de Mishana como representando tres unidades de terreno y no solo dos. Eso se debió a que una parcela de la terraza y una de varillal fueron florísticamente relativamente semejantes entre sí, pero distintas de las otras parcelas. Además, estas dos parcelas compartían algunas características ambientales especiales que originalmente no se habían tomado en cuenta. La parcela de terraza estaba situada al pie de la primera colina con arena blanca, tenía drenaje malo y presentaba en la superficie una capa de arena lavada de arriba. La parcela en la colina de arena blanca tuvo también drenaje impedido, lo que contrastaba con la situación en las otras parcelas de varillal, donde el drenaje era bueno. Por estas razones se unieron las cuatro parcelas caracterizadas con suelo arenoso y drenaje bueno (varillal seco) bajo el nombre Mishana 2, y las dos parcelas caracterizadas con suelo arenoso y drenaje malo (varillal húmedo) bajo el nombre Mishana 3. El resultado de eso fue que finalmente se reconocieron 15 unidades de terreno o sectores donde se hicieron inventario de árboles, y el muestreo en cada sector consistió de dos a cuatro parcelas de tamaño de 25 x 25 m o de 20 x 20 m. La Tabla 7.1 indica la superficie inventariada en cada uno de los sectores.
Dentro de las parcelas, se registraron todos los árboles con troncos más gruesos o iguales de 7,9 cm de circunferencia a la altura del pecho (corresponde a diámetro en la altura del pecho o DAP mayor o igual a 2,5 cm). Se midió la circunferencia del tronco y la altura del árbol, y se recolectó una muestra botánica de cada árbol para la identificación de la especie. Las muestras se obtuvieron usando una tijera telescópica, y en caso de árboles altos se subió hacia la copa del árbol con subidores para llegar a las ramas. Especies fáciles de identificar solo fueron colectadas una vez dentro de cada parcela, pero si había la menor duda sobre la identidad de un árbol se tomó una muestra de éste para su identificación posterior. En todas las parcelas también se muestrearon las pteridófitas y Melastomatáceas de una manera similar que en los transectos (ver abajo).
Tabla 7.1. Los tipos de datos obtenidos en cada uno de los sectores de muestreo. Los códigos de las muestras de suelo están indicados en paréntesis; en el Apéndice 7.4 se presentan los resultados de los análisis de laboratorio para cada una de las muestras. El número de pixels indica cuantos pixels fueron muestreadas para el análisis numérico de la imagen de satélite.
| Pares de variables |
R
|
Pares de variables
|
R
|
Pares de variables
|
R
|
| LLANURA DE fNUNDA06N (COMPLEJO DE ORILLALES) | |||||
| pH vs. arena |
,68
|
Ph vs lLimo
|
-,61
|
PH vs. Arcilla
|
-,77
|
| K-camb. “ “ |
,84
|
K-camb.
|
-,81
|
K-camb.
|
-'81
|
| Sat. Bases “ “ |
,60
|
Sat. bases
|
-,56
|
Sat. bases
|
-,62
|
| C-Org. “ “ |
-,68
|
C-org.
|
,66
|
C-Org. “ “
|
,67
|
| P-disp. “ “ |
-,80
|
P-disp.
|
,85
|
P-disp. “ “
|
,60
|
| Ca-camb. “ “ |
-,82
|
Ca-camb.
|
,81
|
Ca-camb. “ “
|
,75
|
| Mg-camb. “ “ |
-,82
|
Mg-carnb.
|
,76
|
Mg-camb. “ “
|
,86
|
| Suma bases “ “ |
-,82
|
Suma bases
|
,80
|
Suma bases “ “
|
,78
|
| Ac.-camb. “ “ |
-,61
|
Ac.-camb.
|
,56
|
Ac.-camb. “ “
|
,69
|
| CICE “ “ |
-,81
|
CICE
|
,79
|
CICE “ “
|
,78
|
| Sat. Al “ “ |
-,60
|
Sat. Al
|
,56
|
Sat. Al “ “
|
,62
|
| Suma bases “lirno |
,80
|
Suma bases “
|
arcilla -,78
|
Suma bases “arc.+limo
|
,83
|
| CICE “ |
,79
|
CICE “
|
,78
|
CICE “
|
,82
|
| LLANURA DE DESBORDE |
|
||||
| P-disp. C-org. “ |
,70
|
Surna bases
|
-,92
|
|
|
| Ca-camb. “ |
,96
|
Acidez camb.
|
-,94
|
|
|
| Mg-camb. “ |
,95
|
CICE
|
-,93
|
|
|
| Surna bases “ |
,97
|
Sat. Al
|
-,92
|
|
|
| CICE “ |
,97
|
|
|
|
|
| LLANURA DE DESBORDE |
|
|
|
|
|
| K-camb. “ “ |
-,96
|
Sat. bases “ “
|
,92
|
|
|
| Sat. bases “ “ |
-,86
|
|
|
|
|
TRANSECTOS DE COMPOSICIÓN FLORÍSTICA DE PTERIDÓFITAS Y MELASTOMATÁCEAS
En cada uno de los sitios donde se establecieron parcelas para muestrear árboles, con la excepción de Momón, también se estableció un transecto para muestrear pteridófitas y Melastomatáceas con un mayor detalle. Los transectos fueron de 5 metros de ancho y 500 o 1300 metros de largo, dependiendo del nivel de detalle en que se deseaba observar la variación dentro del sitio. Además, se establecieron nueve transectos de 5 m por 500 m para hacer averiguaciones rápidas de la composición florística del bosque en sitios donde no se muestrearon árboles. También se hizo un transecto largo de 2 m por 23 km. (Carbajal), del cual se utilizaron aquí siete secciones de 500 m, una por cada muestreo de suelo que se hizo en los kilómetros 0, 2, 4, 6, 8, 10 y 12.
En algunos transectos se contaron todos los individuos de las pteridófitas y Melastomatáceas y se estimaron sus tamaños, pero en la mayoría sólo se registró la presencia de especies sin información de su abundancia. Para la confirmación posterior de las identificaciones, se colectó en cada sitio por lo menos una muestra botánica de cada especie, y además se colectaron muestras de todos los individuos que no se podían identificar con seguridad en el campo.
Cada transecto tenía su inicio en bosque primario, y el rumbo fue elegido con la ayuda de imágenes de satélite y habitantes de la comunidad más cercana. De esta forma el transecto comprendió bosque primario y no fue interceptado por purmas o chacras. Una vez establecido el rumbo, los transectos avanzaron en línea recta, cruzando tanto bosque alto y maduro como claros recientes, y se impidió los desvíos en las cimas de colinas ó partes fangosas de los valles. De esta manera, sólo los primeros pocos metros de cada transecto se eligieron subjetivamente, y uno puede confiar en que los transectos muestrearon de forma aleatoria y representativa la variación local en la topografía, condiciones edáficas y estados sucesionales del bosque.
Las posiciones geográficas de algunos sitios de muestreo se estimaron midiendo la dirección con brújula y la distancia contando los pasos desde las chacras, purmas, carreteras, vueltas de ríos u otros puntos identificables en la imagen de satélite. En la mayoría de los sitios de muestreo, sin embargo, se midieron las coordenadas geográficas directamente con un GPS portátil (Sistema de Posición Geográfica, modelo Apelco 15). El GPS funciona de tal manera que busca contacto con satélites, y cuando recibe información de por lo menos tres satélites al mismo tiempo, puede determinar su posición (latitud y longitud). En las coordenadas obtenidas puede haber un error hasta de 100-150 m, y por eso es preferible medir las coordenadas en varios puntos para obtener un promedio más confiable. Pero en un bosque alto el dosel impide la comunicación con los satélites, y es relativamente difícil conseguir las coordenadas, excepto en claros relativamente grandes y recientes. Por esta razón, sólo se pudo obtener coordenadas en uno o pocos puntos dentro de cada lugar de muestreo, y hay que aceptar que las coordenadas conseguidas no son exactas.
En el caso de los 17 sitios donde no se muestrearon árboles, el sector de muestreo de las pteridófitas y Melastomatáceas es igual que el transecto establecido en el sitio. Pero en el caso de los 15 sectores donde sí se muestrearon árboles, el sector de muestreo para las pteridófitas y Melastomatáceas incluye tanto las parcelas como el transecto, las que se levantaron dentro de la misma unidad de terreno. En Mishana y 13 de Febrero, donde el transecto cruzó al límite entre tipos diferentes de suelo, el transecto fue dividido en dos partes, y los datos de cada parte se combinaron con los datos de las parcelas que correspondieron al mismo tipo de suelo.
MUESTRAS DE SUELO
En cada sector de muestreo se recolectaron muestras superficiales del suelo. Antes de tomar la muestra se extrajo la hojarasca, raíces y todo el material orgánico existente sobre el suelo mineral. La profundidad de la muestra se midió después de haber quitado este material. Se colectó de 1 a 21 muestras del suelo por sector, normalmente en diversas posiciones topográficas, y se analizó el pH, pérdida por ignición, el contenido de Al, Ca, K, Mg y P, y los porcentajes de diferentes clases granulométricas de cada muestra. Además se hicieron algunas perforaciones más profundas (hasta más que 4 m) describiendo semidetalladamente la variación vertical en el color y en la textura.
En este estudio, para caracterizar los sectores de muestreo, se utilizaron solamente los promedios de los resultados de estas muestras superficiales más algunas otras muestras superficiales colectadas por los pedólogos y geólogos. Todos los métodos y resultados analíticos de esas muestras se presentan en el Apéndice 7.4.
METODOS PARA LOS ANALISIS DE LOS DATOS
IMAGEN DE SATÉLITE
La imagen de satélite Landsat TM que se utilizó en el presente trabajo fue tomada el 23 de Julio de 1993 (véase el Mapa geoecológico, Anexado en este libro). Las diferencias y semejanzas florísticas entre los sectores de muestreo se compararon con los patrones de coloración en la imagen de satélite, tanto en una manera visual como en una manera numérica. En el análisis visual, se observaron los patrones visibles en la imagen y se trató de entenderlos tanto en términos de la composición florística del bosque como en términos de la geología y geomorfología.
Para el análisis numérico, se ubicó en cada sector de muestreo un área alrededor del muestreo florístico (según la longitud del transecto esta área fue desde 24,3 hasta 90 ha, ó entre 270 y 1000 pixels, véase la Tabla 7.1). De esas áreas se extrajeron los valores promedios de reflectancia de luz en seis diferentes longitudes de honda, o sea las bandas 1 a 5 y la banda 7 (véase el recuadro abajo). Las reflectancias promedio fueron utilizadas para caracterizar numéricamente cada uno de los sectores de muestreo. Lamentablemente, en muchos sectores la presencia de nubes 6 una perturbación obvia de actividades humanas impidió la medida confiable de reflectancia del bosque natural. Consecuentemente, para estos análisis sólo se podían utilizar diez de los sectores con muestreo de árboles, y 23 de los sectores con muestreo de pteridófitas y Melastomatáceas. El programa ER-Mapper 5.5 fue utilizado para trabajar digitalmente con la imagen.
| IMÁGENES DE SATÉLITE |
|
La imagen de satélite Landsat TM consiste de células o Pixeles que corresponden a 30 m x 30 m en el campo. Para cada pixel, el satélite registra valores de reflectancia de luz solar en siete áreas diferentes de longitud de honda, llamadas las bandas 1 a 7. Las tres primeras bandas representan longitudes de honda de luz visible: 0,45-0,52 ðmm (banda l); 0,52-0,60 ðmm (banda 2) y 0,63-0,69 ðmm (banda 3). Las bandas 4, 5 y 7 corresponden a luz infrarroja: 0,760,90 ðmm (banda 4); 1,55-1,75 ðmm (banda 5) y 2,08-2,35 ðmm. (banda 7). La banda 6 representa radiación térmica (10,4-11,7 ðmm) y sus pixels son más grandes que los pixels de las otras bandas, 120 m x 120 m. La banda 6 no fue utilizada en el presente trabajo. |
MEDIDAS DE DIFERENCIAS Y SEMEJANZAS ENTRE LOS SECTORES DE MUESTREO
Una idea general en este estudio fue investigar si los cambios en la composición florística están relacionados con otros cambios del medio ambiente. En otras palabras, se trata de ver si los sectores de muestreo que comparten una flora semejante, también son semejantes en términos edáficos, y viceversa. Para este fin, es necesario utilizar algún método exacto para expresar el grado de semejanza ó diferencia entre los sectores de muestreo, tanto en base de la composición florística como en base de las características de los suelos y la imagen de satélite.
En las comparaciones florísticas entre los sectores de muestreo se utilizó el índice de similitud de Jaccard. Para diferencias y semejanzas en términos de las características del suelo o reflectancias de luz (medida por el satélite) se utilizó la distancia euclidiana convertida en un índice de similitud (véase el recuadro abajo). Un índice de similitud consigue valores altos cuando los lugares comparados son semejantes, y valores bajos cuando los lugares son diferentes. Se calculó los valores de los índices entre cada par de lugares de muestreo, y se dispusieron los índices en tablas ó matrices donde se presentan los valores en un orden sistemático (véase un ejemplo en la Tabla 7.8). Todos los cálculos de semejanzas y diferencias, igual que los análisis siguientes, se hicieron con el R-package (Legendre & Vaudor 1991), que es un paquete de programas para análisis multivariante.
| EL ÍNDICE DE JACCARD Y LA DISTANCIA EUCLIDIANA CONVERTIDA |
|
El valor del índice de Jaccard (Legendre & Legendre 1983) puede variar entre 0 y 1, donde 0 significa que los dos sectores no comparten ni una especie, y 1 significa que los dos sectores tienen exactamente las mismas especies. El índice de Jaccard multiplicado por 100 es igual al porcentaje de especies compartidas del número total de especies encontradas en los dos lugares. Sólo se considera la presencia 6 ausencia de las especies en cada lugar, no las abundancias de estas, La distancia euclidiana convertida se calculó en base de la distancia euclidiana. La distancia euclidiana se calcula a base al teorema de Pythagoras. Por ejemplo, en la comparación de los sectores 1 y 2 en base a los promedios de las tres bandas de luz visible (bandas 1, 2 y 3) la fórmula es: distancia euclidiana = "raíz cuadrada" de (b1 size="-2">1-b2 size="-2">2)˛+ (b2 size="-2">1-b2 size="-2">2)˛+ (b3 size="-2">1-b3 size="-2">2)˛ en cual: El valor mínimo de la distancia euclidiana es 0, lo que significa que los dos sectores de muestreo son idénticos considerando las características utilizadas. El valor del índice es siempre más grande cuando los dos sectores son diferentes. Eso representa un comportamiento opuesto al del índice de Jaccard, el cual obtiene valores mayores cuando los dos sectores son similares. Por eso se dice que el índice de Jaccard es un índice de similitud, mientras que la distancia euclidiana es un índice de diferencia. El objetivo en el presente estudio fue investigar, en términos generales, si los sectores que son semejantes a base de un criterio (por ejemplo, composición de especies de pteridófitas) también son semejantes cuando se usa otro criterio (por ejemplo, reflectancias de luz en la imagen de satélite). Para tales comparaciones es preferible utilizar índices con el mismo comportamiento, y eso se obtuvo en este caso con la conversión de los valores de la distancia euclidiana a valores de similitud (DE') en la manera siguiente: DE'= 1- distancia euclidiana La distancia euclidiana convertida (DE') es un índice de similitud comparable al índice de Jaccard. Antes de utilizar los promedios de las características del suelo para calcular las distancias euclidianas convertidas entre los sectores, se estandarizaron los valores de las características del suelo como para tener el mismo promedio (0) y la misma varianza (1) en todas las variables. La estandarización se hizo substrayendo del valor original el promedio de la variable, y después dividiéndolo por la desviación estándar de dicha variable. La estandarización fue necesaria porque las diferentes características del suelo tienen valores absolutos y rangos muy diferentes; por ejemplo el pH varía relativamente poco alrededor del valor 4, mientras que por ejemplo el porcentaje de arena varía de 10 hasta 90. Si uno utiliza los valores originales y junta varias características para calcular la distancia euclidiana, el resultado sería fuertemente dominado por las variables que tienen mucha variación absoluta en sus valores (como el porcentaje de arena). Las variables cuyos valores absolutos son relativamente pequeños y semejantes (como el pH) no afectarían casi en nada al valor del índice. El efecto de la estandarización es darles la misma ponderación a cada variable en el cálculo de la distancia euclidiana convertida. |
VISUALIZACIÓN DE LAS DIFERENCIAS Y SEMEJANZAS ENTRE LOS SECTORES DE MUESTREO
Para visualizar la información de semejanzas y diferencias calculadas (los valores de los índices de similitud) entre los sectores de muestreo, se utilizó la técnica de ordenación de coordenadas principales (Legendre & Legendre 1983). En la ordenación se presentan los sectores de muestreo como puntos en una figura de tal manera que los sectores semejantes según el criterio utilizado aparecen como puntos cercanos entre sí, y los sectores diferentes se sitúan lejanos entre sí (véase el siguiente recuadro).
CORRELACIONES ENTRE LAS MATRICES DE SIMILITUD
La prueba de Mantel (1967) fue utilizada para estudiar matemáticamente si existe correlación linear entre las matrices de similitud. Se aplicó la forma estandarizada de esta prueba (Smouse et al. 1986) que da un valor del coeficiente de correlación entre -1 (correlación negativa completa) y +l (correlación positiva completa). Para estimar la probabilidad del error en el signo del coeficiente de correlación se efectuaron 999 aleatorizaciones (véase el recuadro en la pagina 266).
| LAS MATRICES DE SIMILITUD Y EL MÉTODO DE ORDENACIÓN |
|
Las matrices de similitud presentan, de manera condensada, las relaciones entre los sectores de muestreo, pero sólo considerando los valores del índice es difícil formarse una idea general de cuál es el patrón de las semejanzas y diferencias. Una situación parecida se puede encontrar en guías viales, que presentan cuadros con distancias en kilómetros entre las ciudades principales de un país. Es fácil chequear en tal cuadro cuál es la distancia entre dos ciudades de interés. Pero si uno quiere obtener una idea general sobre la distribución geográfica de las ciudades, es mucho más sencillo verlo en un mapa que en un cuadro de distancias. Uno puede pensar que la tarea de la visualización de los patrones en cualquier matriz de distancia o similitud es como producir un mapa de ciudades en base a las distancias medidas entre ellas. Si la matriz de interés es de similitudes, hay que pensar que primero se convierten los valores de similitud a valores de distancia (distancia = 1- similitud). En el mapa, se dibujan cerca entre sí los lugares con valores de distancia pequeñas, y lejos uno del otro los lugares con distancias grandes. Si tenemos una matriz de distancias entre ciudades, es relativamente fácil construir el mapa, porque sabemos que las ciudades se encuentran prácticamente en un plano de dos dimensiones (latitud y longitud). La tercera dimensión (altitud) sí existe, pero las diferencias en altitud son muy pequeñas en comparación con las distancias horizontales, y consecuentemente no se pierde mucha información aunque se ignore la altitud. En el caso que la matriz de similitud esté basada en, por ejemplo, la composición florística de sitios de muestreo, no hay ninguna razón para pensar que existen sólo dos dimensiones importantes. Además, aunque uno pensaría que dos dimensiones son suficientes para describir los patrones, no podemos predecir que orientaciones van a tener. En realidad, para presentar un mapa exacto de por ejemplo 10 sitios en base a una matriz de similitud, se necesitan 9 dimensiones en un espacio donde cada dimensión forma un ángulo de 90 grados con todas las otras dimensiones. En términos generales, una matriz de n lugares requiere n-1 dimensiones para una presentación completa. Un espacio de más de tres dimensiones ortogonales sobrepasa la capacidad de imaginación, pero matemáticamente es posible construirlo. Existen varios métodos matemáticos para calcular las dimensiones; aquí se utilizó el método "ordenación de coordenadas principales" (Legendre & Legendre 1983). Este método es flexible de tal manera que uno mismo puede elegir el índice de similitud o distancia que sea el más apto para los datos. En el ejemplo de las distancias entre ciudades, se notó que dos dimensiones (latitud y longitud) presentaron más variación que la tercera (altitud), ó sea, eran más importantes. De la misma manera se puede pensar que en un caso de n lugares, algunas de las n-1 dimensiones son más importantes que otras. En efecto, la ordenación de coordenadas principales organiza las dimensiones de tal manera que la primera dimensión es el más importante, y cada una de las dimensiones siguientes es menos importante que la anterior, hasta la última dimensión que es la menos importante. Para una visualización concreta y entendible, que se puede imprimir en papel, lo mejor es elegir las dos primeras dimensiones y dibujar los sitios en un mapa a base de las coordenadas que el programa da para estas dos dimensiones o ejes.
|
| LA PRUEBA DE MANTEL |
|
La visualización por medio de ordenación de los patrones de semejanzas y diferencias entre sectores de muestreo es muy útil cuando se trata de ver si hay relación entre la composición florística y las características ambientales. Si dos ordenaciones de los sectores, una en base a la composición florística y otra en base a alguna característica ambiental, dan posiciones semejantes a los sectores, se puede concluir que la variación de la característica ambiental está correlacionada con la variación de la composición florística. Pero en base a las ordenaciones es difícil de cuantificar el grado de la correlación, evaluar su significado estadístico, o estimar cuáles de las características ambientales son las más altamente correlacionadas con la composición florística. Para obtener respuesta a estas preguntas, se puede hacer un análisis de correlación directamente entre las matrices de similitud. Un método para calcular correlaciones entre matrices es la prueba de Mantel (1967). En esta prueba, la similitud observada (Z) entre dos matrices (X y Y) se calcula como: Z = Eˇ Ij Xij Yˇj donde Xij e Yij corresponden a las similitudes entre los sectores i y j en las dos matrices. El valor observado de Z se compara con el valor esperado de Z, que se obtiene por un método de sorteo al azar. Los valores de similitud en la matriz X se mantienen fijos, mientras en la matriz Y se sortean; después se calcula Z de nuevo. Cuando se repiten el sorteo y el cálculo de Z varias veces, la variación en los valores de Z adquiere una distribución que permite la estimación de la probabilidad de conseguir al azar la similitud observada originalmente. Por ejemplo, si se hacen 99 sorteos y uno de ellos resulta en un valor de Z más extremo que el valor observado originalmente, se tiene en total 2 valores iguales o más extremos que el original de los 99+1 veces de haber calculado Z. Eso corresponde a una significación estadística de p< 2/100, o sea, p4,02. En el presente estudio se hizo el sorteo 999 veces para estimar la significación estadística de la similitud original al nivel de p4,001. Aquí se utilizó una forma estandarizada de Z (Smouse et al. 1986) que siempre está entre +l (correlación positiva perfecta) y -1 (correlación negativa perfecta), y es igual a la correlación linear de Pearson. La estandarización no afecta a las probabilidades asociadas. |
ÁRBOLES
En total se hicieron observaciones de 6253 individuos y 1416 especies de árboles en los 15 sectores donde se muestrearon árboles (Tabla 7.2 y Apéndice 7.1). La acumulación de especies como función del aumento de individuos no demostró ninguna nivelación, aunque lentamente se disminuyó el tacto de la observación de especies nuevas (Figura 7.2). En la riqueza de especies no hubo muchas diferencias entre los sectores de muestreo: en general se encontró alrededor de 200 especies por sector. Es un poco complicado comparar, de manera exacta, la diversidad de especies entre los sectores, porque tanto la superficie muestreada como el número de individuos encontrado variaba entre los sectores. Pero en términos generales se puede decir que en casi todos los sectores se encontró un promedio de más ó menos 2 individuos por especie. Los sectores que más se diferenciaron de los otros son Mishana 2 y Mishana 3, en donde la concentración de especies fue menor.

| Figura 7.2. La acumulación del número de especies de árboles como función del número de individuos en los 15 sectores inventariados. El cálculo está hecho de tal manera que primero se contaron los individuos y el número de especies en uno de los sectores de muestreo, y después se añadió a este listado los individuos de los otros sectores, sector por sector, y cada vez se contó de nuevo el número de individuos y especies. Los sectores se tomaron en cuenta en el siguiente orden: Tarapoto, Gengen, Ex Petroleros, Momón 2, San Antonio 1, 13 de Febrero arcilla, Santa Ana, Nauta, Momón 1, Manití, Panguana, Mishana 1, 13 de Febrero arena, Mishana 3 y Mishana 2. |
Las especies pertenecen a por lo menos 75 familias diferentes; 18 especies quedaron sin identificación de la familia (Tabla 7.3). Las familias más ricas en especies son, en general, las que más individuos tienen, aunque hay algunas excepciones a esta regla. Las más notables son Myristicaceae, Violaceae y Rutaceae que tienen mucho más individuos que su riqueza en especies sugiere. Por otro lado, Lauraceae, Myrtaceae y Elaeocarpaceae son familias bien ricas en especies, pero son representadas por relativamente pocos individuos.
Ninguna de las especies fue observada en todos los sectores de muestreo, y la mayoría de las especies (811 ó 57,7 %) sólo fueron observadas en un sector (Figura 7.3A). Apenas 30 especies (2,1 %) fueron observadas en más de 50 % de los sectores (Figura 7.3A, Tabla 7.4). Las cuatro especies más abundantes tuvieron más de 60 individuos cada una, pero las 625 especies menos abundantes presentaban un solo individuo cada una (Figura 7.313). Las 32 especies más abundantes representaron 20 % de todos los individuos (Tabla 7.5). Es notable que tanto en términos de frecuencia como en términos de abundancia hay varias especies de Myristicaceae que presentan cifras elevadas. También en las familias Burseraceae, Euphorbiaceae y Violaceae se encuentran varias especies frecuentes y/o abundantes.
Tabla 7.2. Area inventariada, número de individuos y número de especies de árboles encontrados en los 15 sectores de muestreo de árboles. En la columna del número de especies se indica en paréntesis el número promedio de individuos por especie.
| SECTOR | Área inventariada (ha) | Número de iondividuos | Número de especies (individuos por especie) |
| Tarapoto |
0,16
|
413
|
234(1,8)
|
| Gengen |
0,16
|
435
|
220(2,0)
|
| Ex Petroleros |
0,16
|
400
|
240(1,7)
|
| San Antonio 1 |
0,16
|
546
|
275(2,0)
|
| Momón 2 |
0,125
|
330
|
165(2,0)
|
| 13 de Febrero arcilla |
0,08
|
150
|
114(1,3)
|
| Santa Ana |
0,16
|
368
|
234(1,6)
|
| Nauta |
0,16
|
447
|
245(1,8)
|
| Mishana 1 |
0,1875
|
532
|
195(2,7)
|
| Momón 1 |
0,125
|
297
|
145(2,0)
|
| Maniti |
0,16
|
459
|
214(2,1)
|
| Panguana 1 |
0,16
|
437
|
240(1,8)
|
| 13 de Febrero arena |
0,08
|
203
|
135(1,5)
|
| Mishana 3 |
0,125
|
435
|
132(3,3)
|
| Mishana 2 |
0,25
|
801
|
159(5,0)
|
| Total |
22,525
|
6253
|
1416(4,4)
|
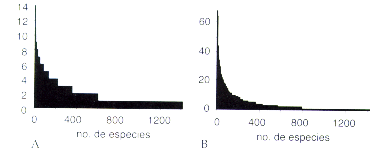 | Figura 7.3. Diagramas de especies de árboles en los 15 sectores inventariados. A. Distribución de las frecuencias entre las especies. B. Distribución de la abundancia entre las especies. |
Se nota que las escalas en los ejes verticales son diferentes: la frecuencia no puede sobrepasar el valor 15, lo que significa que la especie fue encontrada en todos los sectores de muestreo, mientras que el número de individuos no tiene un valor máximo teórico. Cada unidad en el eje horizontal representa una especie, y las especies están organizadas de izquierda a derecha según disminuya su frecuencia (A) o número de individuos (B).
Tabla 7.3. Número y porcentajes de especies e individuos por familia entre los árboles observados en los 15 sectores de muestreo.
| SECTOR |
Área inventariada (ha)
|
Número de iondividuos
|
Número de especies (individuos por especie)
|
| Tarapoto |
0,16
|
413
|
234(1,8)
|
| Gengen |
0,16
|
435
|
220(2,0)
|
| Ex Petroleros |
0,16
|
400
|
240(1,7)
|
| San Antonio 1 |
0,16
|
546
|
275(2,0)
|
| Momón 2 |
0,125
|
330
|
165(2,0)
|
| 13 de Febrero arcilla |
0,08
|
150
|
114(1,3)
|
| Santa Ana |
0,16
|
368
|
234(1,6)
|
| Nauta |
0,16
|
447
|
245(1,8)
|
| Mishana 1 |
0,1875
|
532
|
195(2,7)
|
| Momón 1 |
0,125
|
297
|
145(2,0)
|
| Maniti |
0,16
|
459
|
214(2,1)
|
| Panguana 1 |
0,16
|
437
|
240(1,8)
|
| 13 de Febrero arena |
0,08
|
203
|
135(1,5)
|
| Mishana 3 |
0,125
|
435
|
132(3,3)
|
| Mishana 2 |
0,25
|
801
|
159(5,0)
|
| Total |
22,525
|
6253
|
1416(4,4)
|
| Familia | Especie | Frecuencia | Número de Individuos | %de Individuos |
| Myristicaceae | Iryanthera macrophylla |
14
|
34
|
0,54
|
| Myristicaceae | Virola calophylla |
14
|
68
|
1,09
|
| Olacaceae | Tetrastylidium peruvianum |
14
|
34
|
0,54
|
| Myristicaceae | Iryanthera cf. paraensis |
12
|
37
|
0,59
|
| Boraginaceae | Cordia nodosa |
11
|
24
|
0,38
|
| Cecropiaecae | Pourouma minor |
11
|
16
|
0,26
|
| Nyctaginaceae | Neea cf. verticíllata |
11
|
55
|
0,88
|
| Annonaceae | Unonopsis stipitata |
10
|
34
|
0,54
|
| Burseraceae | Protium nodulosum |
lo
|
28
|
0,45
|
| Burseraceae | Protium opacum |
10
|
22
|
0,35
|
| Euphorbiaceae | Nealchornea yapurensis |
10
|
30
|
0,48
|
| Annonaecae | Guatteria megalaphylla |
9
|
24
|
0,38
|
| Burseraceae | Protium grandifollum |
9
|
17
|
0,27
|
| Euphorbiaceae | Conceveiba rhytidocarpa |
9
|
22
|
0,35
|
| Melastomataceae | Tococa guianensis |
9
|
31
|
0,50
|
| Moraceae | Naucleopsis imitans |
9
|
18
|
0,29
|
| Moraceae | Pseudolmedia laevigata |
9
|
18
|
0,29
|
| Myristicaceae | Virola pavonis |
9
|
62
|
0,99
|
| Myrtaceae | Eugenia florida |
9
|
18
|
0,29
|
| Arecaceae | Oenocarpus bataua |
8
|
41
|
0,66
|
| Acercare | Socratea exorrizha |
8
|
18
|
0,29
|
| Burseraceae | Protium apiculatum |
8
|
38
|
0,61
|
| Flacourtiaceae | Casearia javitensis |
8
|
18
|
0,29
|
| Lecythidaceae | Eschweilera cf. coriacea |
8
|
29
|
0,46
|
| Lecythidaceae | Eschweilera tessmannii |
8
|
16
|
0,26
|
| Leguminosae | Inga tenuistipula |
8
|
17
|
0,27
|
| Moraceae | Perebea xanthochyma |
8
|
16
|
0,26
|
| Myristicaceae | Iryanthera juruensis |
8
|
24
|
0,38
|
| Sterculiaceae | Theobroma subincanum |
8
|
22
|
0,35
|
| Violaceae | Leonia glycicarpa |
8
|
19
|
0,30
|
Melastomatáceas
En los 32 sectores de estudio donde se hizo inventario de las Melastornatáceas, se observaron en total 134 especies de esta familia, representando por lo menos 13 géneros diferentes; dos especies no fueron identificadas hasta el género (Apéndice 7.2). En varios de los sectores se registró solamente la presencia de las especies sin contar los individuos, y por lo tanto no es posible dar cifras exactas del número de individuos observados. Pero se puede calcular un promedio de individuos de Melastomatáceas por área, utilizando los sectores donde sí se contaron los individuos: en un área de 6,25 ha se observaron 9507 individuos de Melastomatáceas, lo cual resulta en un promedio de 1521 individuos por hectárea. El área total de los inventarios de las Melastomatáceas en los 32 sectores llega a 10,46 ha, así que se puede estimar que a los resultados del presente estudio han contribuido unos 16 000 individuos de Melastomatáceas.
Tabla 7.5. Las 32 especies de árboles más abundantes (con el mayor número de individuos) en los 15 sectores inventariados.
| FAMILIA | Especie |
Número deindividuos
|
% de individuos
|
Frecuencia
|
| Myristicaceae | Virola calophylla |
68
|
1,09
|
14
|
| Violaceae | Rinorea racemosa |
68
|
1,09
|
6
|
| Rutaceae | Ravenia biramosa |
64
|
1,02
|
1
|
| Myristicaceae | Virola pavonis |
62
|
0,99
|
9
|
| Nyctaginaceae | Neea cf. verticillata |
55
|
0,88
|
11
|
| Euphorbiaceae | Mabea speciosa |
53
|
0,85
|
7
|
| Sapotaceae | Chrysaphyllum sanguinolentum |
52
|
0,83
|
5
|
| Leguminosae | Macrolobium microcalyx |
44
|
0,70
|
2
|
| Acercare | Oenocarpus bataua |
41
|
0,66
|
8
|
| Monimiaceae | Siparuna guianensis |
41
|
0,66
|
7
|
| Violaceae | Rinorea flavescens |
39
|
0,62
|
4
|
| Burseraceae | Protium apiculatum |
38
|
0,61
|
8
|
| Euphorbiaceae | Senefeldera macrophylla |
37
|
0,59
|
4
|
| Lecythidaceae | Eschweilera cf. andina |
37
|
0,59
|
6
|
| Myristicaceae | Iryanthera cf. paraensis |
37
|
0,59
|
12
|
| Leguminosae | Macrolobium limbatum |
36
|
0,58
|
6
|
| Annonaceae | Unonopsis stipitata |
34
|
0,54
|
10
|
| Myristicaceae | Iryanthera macrophylla |
34
|
0,54
|
14
|
| Olacaceae | Tetrastylidium peruvianum |
34
|
0,54
|
14
|
| Myristicaceae | Otoba gIycycarpa |
33
|
0,53
|
6
|
| Lecythidaceae | Eschweilera sp. 1 |
32
|
0,51
|
5
|
| Annonaceae | Diclinanona tessmannii |
31
|
0,50
|
7
|
| Bombacaceae | Rhodognaphalopsis brevipes |
31
|
0,50
|
2
|
| Melastomataceae | Tococa guianensis |
31
|
0,50
|
9
|
| Euphorbiaceae | Nealchornea yapurensis |
30
|
0,48
|
10
|
| Euphorbiaceae | Mabea sp. 1 |
29
|
0,46
|
3
|
| Lecythidaceae | Eschweilera cf. coriacea |
29
|
0,46
|
8
|
| Leguminosae | Pithecellobium basijugum |
29
|
0,46
|
7
|
| Myristicaceae | Iryanthera tricornis |
29
|
0,46
|
5
|
| Burseraceae | Protium nodulosum |
28
|
0,45
|
10
|
| Euphorbiaceae | Senefeldera inclinata |
28
|
0,45
|
5
|
| Myristicaceae | Iryanthera polyneura |
27
|
0,43
|
6
|
Seis especies (4,4 %) de las Melastomatáceas fueron encontradas en más del 50 % de los sectores de estudio, mientras que 32 especies (23,7 %) se observaron sólo en un sector (Figura 7.4). Las especies más frecuentemente encontradas están en el listado de la Tabla 7.6.
Tabla 7.6. Las 20 especies más frecuentes de las Melastomatáceas en los sectores de estudio.
| ESPECIE(número de sectores), |
Frecuencia (% de ectores)
|
Frecuencia
|
| Maieta guianensis |
24
|
75
|
| Tococa ulei |
23
|
72
|
| Miconia tomentosa |
22
|
69
|
| Miconia klugii |
19
|
59
|
| Tococa guianensis |
19
|
59
|
| Miconia prasina |
16
|
50
|
| Miconia nervosa |
15
|
47
|
| Clidemia epiphytica |
14
|
44
|
| Miconia punctata |
14
|
44
|
| Miconia carassana |
13
|
41
|
| Clidemia longifolia |
12
|
38
|
| Leandra longicoma |
12
|
38
|
| Leandra macdanielii |
12
|
38
|
| Ossaea araneifera |
12
|
38
|
| Miconia serrulata |
11
|
34
|
| Miconia cf. subspicata |
11
|
34
|
| Miconia traillii |
11
|
34
|
| Ossaea boliviensis |
11
|
34
|
| Miconia sp. 55 |
10
|
31
|
| Miconia tetrasperma |
10
|
31
|
| Tococa caquetana |
10
|
31
|
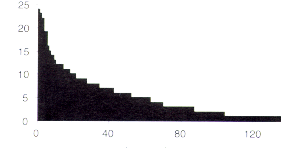 |
Figura 7.4. Distribución de frecuencias entre las especies de Melastomatáceas en los 32 sectores inventariados. Cada unidad en el eje horizontal representa una especie, y las especies están organizadas de izquierda a derecha según la disminución de la frecuencia. La frecuencia máxima, 32, significaría que la especie se encontró en todos los sectores de estudio. |
Pteridófitas
En los 32 sectores de estudio donde se hizo el inventario de las pteridófitas, se observaron en total 152 especies de pteridófitas que representaban 39 géneros diferentes (Apéndice 7.3). En varios de los sectores se registró solamente la presencia de las especies sin contar los individuos, y por lo tanto no es posible dar cifras exactas del número de individuos observados. Pero se puede calcular un promedio de individuos de pteridófitas por área, utilizando los sectores donde sí se contaron los individuos: en un área de 6,25 ha se observaron 47 229 individuos de pteridófitas, lo cual resulta en un promedio de 7557 individuos por hectárea. El área total de los inventarios de las pteridófitas en los 32 sectores totaliza 10,46 ha. Entonces se puede estimar que a los resultados del presente estudio han contribuido unos 80 000 individuos de pteridófitas.
Las frecuencias de las especies de pteridófitas son bastante parecidas a las frecuencias de las Melastomatáceas, o un poco más altas. Catorce especies de pteridófitas (9,2 %) se encontraron en más del 50 % de los sectores de estudio, mientras 38 especies (21,1 %) se observaron sólo en un sector (Figura 7.5). Las especies más comúnmente encontradas se incluyen en la lista de la Tabla 7.7.
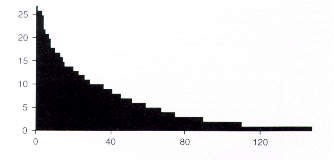 |
Figura 7.5. Distribución de frecuencias entre las especies de pteridófitas en los 32 sectores inventariados. Cada unidad en el eje horizontal representa una especie, y las especies están organizadas de izquierda a derecha según la disminución de la frecuencia. La frecuencia máxima, 32, significaría que la especie fue encontrada en todos los sectores estudiados. |
Tabla 7.7. Las 20 especies mas frecuentes de pteridófitas en los 32 sectores de estudio.
| ESPECIE |
Frecuencia (# de sectores)
|
Frecuencia (% de sectores)
|
| Cyathea lasiosora |
27
|
84
|
| Lomariopsis nigropaleata |
26
|
81
|
| Trichomanes pinnatum s.l. |
26
|
81
|
| Nephrolepis rivularis |
25
|
78
|
| Metaxya rostrata |
22
|
69
|
| Cyc1odium meniscioides |
21
|
66
|
| Polybotrya osmundacea |
21
|
66
|
| Saccoloma inaequale |
20
|
63
|
| Asplenium serratum |
18
|
56
|
| Salpichlaena volubilis |
18
|
56
|
| Adiantum sp. 3 ó Adiantum sp. 4 |
17
|
53
|
| Lindsaea divaricata |
17
|
53
|
| Polybotrya pubens |
17
|
53
|
| Lomagramma guianensis |
16
|
50
|
| Po1ybotrya caudata |
15
|
47
|
| Adiantum humile |
14
|
44
|
| Antrophyum cayennense ó A. guianense |
14
|
44
|
| Cyathea pungens |
14
|
44
|
| Microgramma thurnii |
14
|
44
|
| Schizaea elegans |
14
|
44
|
PATRONES FLORÍSTICOS Y AMBIENTALES
Las 49 parcelas con inventarios de árboles
El muestreo de los árboles consistió de 49 parcelas de un tamaño de 20 x 20 m o de 25 x 25 m, las que se distribuyeron entre los 15 sectores de muestreo de tal manera que cada sector incluyó entre dos y cuatro parcelas. La Figura 7.6 presenta una ordenación que muestra del patrón de semejanzas entre las 49 parcelas, calculadas en base a la composición florística de árboles. En la ordenación se puede apreciar que las parcelas levantadas en el mismo sector, los cuales fueron definidos geográficamente o en base a una diferencia marcada en el suelo, son florísticamente relativamente cercanas entre si. Sólo en el caso de los sectores de Mishana había discrepancia entre la clasificación original del terreno y los patrones florísticos, ya que originalmente sólo se habían reconocido dos unidades de terreno en Mishana (Mishana 1 y Mishana 2; las parcelas marcadas Mishana 3 en Figura 7.6 fueron originalmente divididas entre los otros dos sectores, ya que una se localizó en la planicie de arena blanca y otra en la zona límite entre la terraza y la planicie de arena blanca). Para explicar esta discrepancia, se volvió a averiguar las características del terreno, y se observó que en realidad las dos parcelas que no conformaban al patrón florístico previsto, tenían una combinación única de suelo arenoso y drenaje malo. Por lo tanto se decidió reconocer un tercer sector en Mishana.
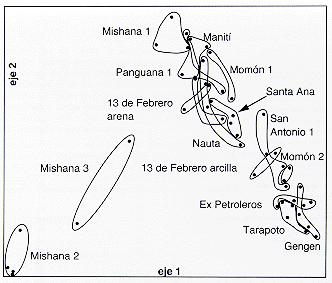 |
Figura 7.6. Ordenación de coordenadas principales basadas en las composiciones florísticas de árboles (Indice de Jaccard). Cada punto representa una de las 49 parcelas de tamaño de 20 x 20m o de 25 x 25 m. Las parcelas que representan el mismo sector de estudios están incluidas dentro de la misma línea. |
Los 15 sectores con inventarios de árboles
Las Tablas 7.8, 7.9 y 7.10 representan las matrices de similitud (utilizando los porcentajes de especies compartidas) entre los 15 sectores de muestreo según los tres grupos de plantas observadas (árboles, Melastomatáceas y pteridófitas). Se puede observar que los porcentajes de especies compartidas tienen rangos diferentes en los tres grupos de plantas: con árboles que varían de 3 á 35 %, con Melastomatáceas que varían de 0 á 60 %, y con pteridófitas que varían de 0 á 54 %. Las ordenaciones basadas en estos porcentajes (Figuras 7.7A-C) muestran los patrones generales de semejanzas y diferencias florísticas entre los sectores. La Figura 7.7D muestra una ordenación basada en una combinación de varias características de los suelos (pH, LO, porcentaje de arena, y los contenidos de Al, Ca, K, Mg y Na).
Tabla 7.8. Porcentaje de especies de árboles compartidos entre todos los pares de los 15 sectores de muestreo.
| . | Gen | ExP | Mo 2 | Ant | F arc | Ana | Nau | Mo 1 | Man | Pang | Mish | F are | Mi 3 | Mi 2 |
| Tarapoto |
33
|
29
|
17
|
23
|
16
|
14
|
13
|
12
|
12
|
11
|
8
|
5
|
5
|
3
|
| Gengen |
|
34
|
30
|
27
|
19
|
18
|
19
|
15
|
18
|
14
|
10
|
10
|
9
|
6
|
| Ex Petroleros |
|
|
25
|
32
|
23
|
21
|
19
|
13
|
15
|
13
|
11
|
12
|
7
|
4
|
| Momón 2 |
|
|
|
25
|
18
|
21
|
15
|
23
|
17
|
13
|
20
|
11
|
13
|
9
|
| San Antonio 1 |
|
|
|
|
24
|
26
|
25
|
22
|
21
|
18
|
20
|
17
|
7
|
7
|
| 13 de Feb. arcilla |
|
|
|
|
17
|
17
|
16
|
15
|
14
|
16
|
18
|
9
|
5
|
|
| Santa Ana |
|
|
|
|
|
|
24
|
19
|
25
|
20
|
19
|
14
|
10
|
10
|
| Nauta |
|
|
|
|
|
|
|
23
|
24
|
26
|
27
|
22
|
14
|
14
|
| Momón 1 |
|
|
|
|
|
|
|
|
23
|
20
|
27
|
18
|
11
|
8
|
| Maniti |
|
|
|
|
|
|
|
|
|
28
|
29
|
19
|
19
|
12
|
| Panguana 1 |
|
|
|
|
|
|
|
|
|
|
29
|
19
|
11
|
10
|
| Mishana |
|
|
|
|
|
|
|
|
|
|
|
24
|
22
|
16
|
| 13 de Feb. arena |
|
|
|
|
|
|
|
|
|
|
|
|
17
|
14
|
| Mishana 3 |
|
|
|
|
|
|
|
|
|
|
|
|
|
35
|
Tabla 7.9. Porcentaje de especies de Melastomatáceas compartidas entre todos los pares de los 15 sectores de muestreo.
| . | Gen | Ex P | Mo 2 | Ant | F arc | : Ana | Nau | Mo 1 | Man | Pang | Mish | F are | Mi3 | Mi2 |
| Tarapoto |
59
|
41
|
6
|
22
|
10
|
11
|
5
|
3
|
5
|
3
|
2
|
7
|
0
|
0
|
| Gengen |
|
55
|
14
|
32
|
17
|
16
|
8
|
5
|
7
|
8
|
5
|
15
|
2
|
0
|
| Ex Petroleros |
|
|
12
|
35
|
19
|
23
|
8
|
8
|
5
|
10
|
8
|
17
|
3
|
0
|
| Momón 2 |
|
|
|
9
|
13
|
10
|
10
|
20
|
8
|
10
|
10
|
12
|
12
|
9
|
| San Antonio 1 |
|
|
|
|
23
|
26
|
14
|
9
|
12
|
15
|
10
|
20
|
6
|
2
|
| 13 de Feb. arcilla |
|
|
|
|
32
|
14
|
19
|
22
|
17
|
23
|
60
|
24
|
14
|
|
| Santa Ana |
|
|
|
|
|
|
29
|
22
|
37
|
31
|
22
|
27
|
12
|
6
|
| Nauta |
|
|
|
|
|
|
|
7
|
25
|
28
|
26
|
14
|
17
|
5
|
| Montón 1 |
|
|
|
|
|
|
|
|
14
|
13
|
9
|
25
|
5
|
8
|
| Maniti |
|
|
|
|
|
|
|
|
|
37
|
26
|
27
|
23
|
9
|
| Panguana 1 |
|
|
|
|
|
|
|
|
|
|
22
|
22
|
24
|
14
|
| Mishana |
|
|
|
|
|
|
|
|
|
|
|
28
|
28
|
7
|
| 13 de Feb. arena |
|
|
|
|
|
|
|
|
|
|
|
|
24
|
14
|
| Mishana 3 |
|
|
|
|
|
|
|
|
|
|
|
|
|
25
|
Tabla 7.10. Porcentaje de especies de pteridófitas compartidas entre todos los pares de los 15 sectores de muestreo.
| . | Gen | Ex P | Mo 2 | Ant | F arc | Ana | Nau | Mo 1 | Man | Pan | Mis | F are | Mi 3 | Mi2 |
| Tarapoto |
54
|
52
|
30
|
36
|
21
|
21
|
17
|
13
|
14
|
23
|
8
|
8
|
2
|
0
|
| Gengen |
|
53
|
32
|
43
|
23
|
29
|
19
|
16
|
18
|
23
|
9
|
9
|
5
|
0
|
| Ex Petroleros |
|
|
29
|
42
|
27
|
30
|
27
|
20
|
24
|
26
|
16
|
10
|
8
|
2
|
| Momon2 |
|
|
|
30
|
29
|
28
|
28
|
33
|
32
|
22
|
20
|
21
|
12
|
3
|
| San Antonio 1 |
|
|
|
|
39
|
47
|
34
|
25
|
29
|
39
|
19
|
15
|
13
|
2
|
| 13 de Feb. arcilla |
|
|
|
|
|
39
|
33
|
39
|
37
|
35
|
30
|
19
|
15
|
5
|
| Santa Ana |
|
|
|
|
|
|
49
|
31
|
44
|
37
|
31
|
18
|
17
|
5
|
| Nauta |
|
|
|
|
|
|
|
38
|
49
|
31
|
41
|
21
|
17
|
4
|
| Momeen 1 |
|
|
|
|
|
|
|
|
44
|
39
|
41
|
27
|
21
|
6
|
| Maniti |
|
|
|
|
|
|
|
|
|
40
|
41
|
23
|
19
|
9
|
| Panguana 1 |
|
|
|
|
|
|
|
|
|
|
39
|
16
|
18
|
4
|
| Mishana |
|
|
|
|
|
|
|
|
|
|
|
27
|
28
|
3
|
| 13 de Feb. arena |
|
|
|
|
|
|
|
|
|
|
|
|
17
|
9
|
| Mishana 3 |
|
|
|
|
|
|
|
|
|
|
|
|
|
23
|
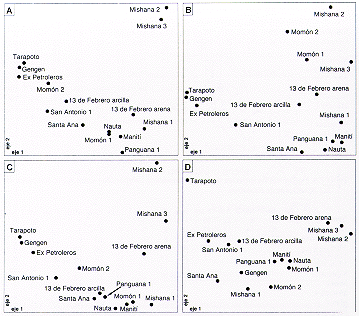 |
Figura 7.7. Ordenaciones de coordenadas principales de los 15 sectores en estudio. A. La composición florística de árboles (índice de Jaccard). B. la composición florística de las Melastomatáceas (índice de Jaccard). C. la composición florística de las pteridófitas (índice de Jaccard). D. características del suelo (distancia euclidiana en base a pH, LO, porcentaje de arena, y contenido de Al, Ca, K, Mg y Na). |
Se puede observar que cada una de las ordenaciones presenta un patrón generalmente semejante, y que especialmente las ordenaciones florísticas son parecidas entre si. Los sectores de muestreo forman una nube curvada, estando en un extremo Tarapoto, Ex Petroleros y Gengen, y en el otro extremo se encuentran Mishana 2 y Mishana 3. También en las partes centrales de las ordenaciones se pueden distinguir otros dos grupos de sectores de muestreo. Casi siempre Momón 2, San Antonio 1, 13 de Febrero arcilla y Santa Ana están en el lado de Tarapoto, mientras Maniti, Panguana 1, Nauta, Mishana 1, Momón 1 y 13 de Febrero arena quedan más cerca de Mishana 2. Las excepciones más evidentes de esta regla son Momón 1 y Momón 2 en el caso de las Melastomatáceas, y Gengen y Momón 2 en el caso de los suelos.
Las características del suelo en los sectores de estudio (Tabla 7.11) sugieren que las posiciones de los sectores en la ordenación (Figura 7.7D) corresponden más que todo a las diferencias en textura y al contenido de cationes en el suelo. Tarapoto y los sectores cercanos en la ordenación tienen porcentaje bajo de arena pero contenido relativamente alto de cationes, y hacia Mishana 2 y Mishana 3 crece la proporción de arena y disminuye el contenido de cationes.
Tabla 7.11. Promedios de los valores de características diferentes del suelo en los 32 sectores de muestreo (en cmolckg-1 ). Las dos columnas separadas para fósforo refieren a los dos métodos diferentes y no directamente comparables que se han utilizado para su análisis (véase Apéndice 7.4). En los 15 sectores marcados con "A% se ha hecho inventarios de árboles, Melastomatáceas y pteridófitas; en los otros sectores sólo se ha hecho inventario de Melastomatáceas y pteridófitas.
| Sector | K | Ca | Mg | Na | Al | pH |
LOI % |
P-Bray | P | Arena | |
| Palo Seco 1 |
0,19
|
8,95
|
0,98
|
0,02
|
3,31
|
4,09
|
7,44
|
10
|
|
11,3
|
|
| Tarapoto | A |
0,2
|
9,17
|
1,3
|
0,02
|
3,43
|
3,97
|
6,8
|
5,67
|
|
10,0
|
| 7 de Julio |
0,15
|
6,67
|
0,32
|
0,02
|
1,62
|
4,18
|
5,73
|
2,52
|
|
20
|
|
| Gengen | A |
0,12
|
0,46
|
0,11
|
0,03
|
8,37
|
3,84
|
5,93
|
|
67,6
|
14,1
|
| Libertad Agraria km. 8 |
0,21
|
4,94
|
0,57
|
0,02
|
6,28
|
3,75
|
7,68
|
2,73
|
|
10,3
|
|
| Ex Petroleros | A |
0,17
|
1,65
|
0,48
|
0,03
|
7,66
|
3,64
|
5,92
|
5,64
|
|
16,1
|
| San Antonio 1 | A |
0,16
|
0,76
|
0,38
|
0,03
|
5,98
|
3,64
|
6,49
|
5,96
|
|
40,5
|
| Carbajal km0 |
0,17
|
1,07
|
0,41
|
0,01
|
7,04
|
3,5
|
6,9
|
8,65
|
|
7
|
|
| Carbajal km2 |
0,15
|
0,38
|
0,26
|
0,04
|
10,4
|
3,58
|
6,37
|
5,92
|
|
9,5
|
|
| Palo Seco 2 |
0,03
|
0,05
|
0,04
|
0
|
0,11
|
2,64
|
3,49
|
6,72
|
|
91
|
|
| Momón 2 | A |
0,04
|
0
|
0,02
|
0,01
|
8,04
|
3,93
|
5,52
|
|
8,09
|
10,3
|
| 13 de Febrero arcilla | A |
0,18
|
0,94
|
0,27
|
0,02
|
4,85
|
3,65
|
5,79
|
1,35
|
|
36,4
|
| Sta. Ana | A |
0,18
|
0,13
|
0,19
|
0,02
|
10
|
3,56
|
8,03
|
1,49
|
|
10,8
|
| Carbajal km4 |
0,12
|
0,15
|
0,17
|
0,03
|
7,04
|
3,78
|
7,24
|
1,16
|
|
29,5
|
|
| Carbajal km8 |
0,08
|
0,83
|
0,28
|
0,03
|
3,41
|
3,88
|
4,7
|
1,72
|
|
50
|
|
| Carbajal kml0 |
0,09
|
0,32
|
0,11
|
0,02
|
4,25
|
3,76
|
4,4
|
1,65
|
|
42,5
|
|
| Carbajal kml2 |
0,11
|
0,08
|
0,14
|
0,03
|
6,91
|
3,58
|
5,99
|
1,51
|
|
36
|
|
| San Antonio 2 |
0,11
|
0,12
|
0,13
|
0,02
|
5,67
|
3,67
|
5,42
|
3,67
|
|
38,2
|
|
| Nauta | A |
0,06
|
0,01
|
0,06
|
0,01
|
3,88
|
3,9
|
5,48
|
|
26,1
|
49,8
|
| Mishana 1 | A |
0,1
|
0,14
|
0,06
|
0,01
|
10,3
|
3,96
|
7,02
|
|
22,8
|
6,3
|
| Momón 1 | A |
0,04
|
0,03
|
0,04
|
0,02
|
4,59
|
4,02
|
3,92
|
|
24,3
|
23,5
|
| Maniti | A |
0,08
|
0,02
|
0,09
|
0,01
|
3,7
|
3,85
|
6,73
|
|
66,9
|
54,6
|
| Panguana 1 | A |
0,05
|
0,04
|
0,06
|
0,04
|
3,85
|
3,91
|
4,34
|
0,81
|
|
42,2
|
| Panguana 2 |
0,04
|
0,05
|
0,04
|
0,01
|
2,72
|
3,93
|
2,98
|
1,24
|
|
27,3
|
|
| Libertad Agraria km. 3 |
0,1
|
0,06
|
0,1
|
0,01
|
5,28
|
3,89
|
7,29
|
1,66
|
|
31,7
|
|
| San Pedro |
0,07
|
0,02
|
0,07
|
0,01
|
5,26
|
3,8
|
5,6
|
2,03
|
|
46,3
|
|
| 13 de Febrero arena | A |
0,06
|
0
|
0,06
|
0,01
|
0,75
|
3,65
|
1,86
|
3,7
|
|
80
|
| San Gerardo |
0,02
|
0,11
|
0,05
|
0,12
|
0,22
|
3,4
|
|
2,51
|
|
90,5
|
|
| Carbajal km6 |
0,27
|
0,19
|
0,44
|
0,07
|
0,41
|
2,39
|
34,7
|
5,32
|
|
53
|
|
| Pto. Almendras |
0,09
|
0,05
|
0,07
|
0,01
|
0,58
|
3,22
|
3,18
|
6,69
|
|
85,3
|
|
| Mishana 3 | A |
0,01
|
0,01
|
0
|
0,01
|
0,63
|
3,71
|
1,29
|
|
5,13
|
77,1
|
| Mishana 2 | A |
0,01
|
0
|
0
|
0
|
1,29
|
4,29
|
1,36
|
|
10,6
|
71,4
|
Según las ordenaciones en la Figura 7.7, se pueden dividir los sectores de estudio en 4 grupos distintos, y es interesante tratar de identificar las especies que caracterizan algún o algunos de estos grupos. La Tabla 7.12 presenta algunas de dichas especies, elegidas entre las especies más comunes de árboles que se han podido identificar hasta la especie misma (véase Apéndice 7.1). Al interpretar la Tabla 7.12 hay que tomar en cuenta que la preferencia indicada a cierta especie significa que según las observaciones de este estudio, hay claramente una probabilidad más alta de encontrar la especie en los lugares indicados, pero eso no significa necesariamente que la especie no crece en otras partes.
Tabla 7.12. Preferencias ecológicas de algunas especies comunes de árboles. Los cuatro círculos representan la gradiente edáfica desde suelos arcillosos con contenido elevado de cationes a la izquierda hacia suelos arenosos con contenido bajo de cationes a la derecha. Los círculos negros significan que las especies mencionadas al costado han sido observadas en la parte respectiva de la gradiente; los círculos blancos significan lo contrario.
| Suelo preferido | Especies de árboles |
| ©OOO | Croton tessmannii, Otoba glycycarpa, Calyptranthes longifólia,Eugenia discreta, Coussarea megalocarpa |
| ©©OO | Tetragastris panamensis, Carpotroche longifolia, Eschweilera cf.coriacea, Perebea xanthochyma, Warscewiczia coccinea, Theobroma obovatum |
| ©©©O | Unonopsis stipitata, Socratea exorrhiza, Protium apiculatum,Protium opacum, Pourouma minor, Conceveiba rhytidocarpa,Nealchornea yapurensis, Eschweilera tessmannii, Inga tenuistipula,Siparuna guianensis, Theobroma subincanum, Leonia glycicarpa |
| O©OO | Lindackeria paludosa |
| O©©O | Senefeldera inclinata, Senefeldera macrophylla, Rinorea flavescens,Rinorea racemosa |
| OO©O | Oenocarpus bataua, Sloanea tuerckheimii, Eschweilera cf. andina,Pithecellobium basijugum, Iryanthera polyneura, Iryanthera tricornis |
| O©©© | Protium grandifolium, Mabea speciosa, Tococa guianensis, Virolapavonis |
| OO©© | Diclinanona tessmannii, Chrysophyllum sanguinolentum |
| OOO© | Aspidosperma exelsum, Dendropanax umbellatus,Rhodognaphalopsis brevipes, Macrolobium limbatum, Macrolobiummicrocalyx, Tachigali paniculata, Ravenia biramosa, Mataybainelegans |
La posición de los sectores con arena blanca (Mishana 2 y Mishana 3) en la ordenación con árboles (Figura 7.7A) sugiere que tienen una composición florística muy especial, y realmente existen varias especies de árboles que solamente se han encontrado en bosques de arena blanca. La Tabla 7.12 menciona para el grupo no. 4 ocho especies que tienen una preferencia ecológica muy clara (Apéndice 7.1).
Además, hay una buena cantidad de especies comunes que crecen en cualquier otro lugar, pero no en arena blanca. No todos los grupos de sectores de inventarío están tan bien definidos. Por ejemplo, de las especies encontradas en el grupo número 2 (Momón 2, San Antonio 1, 13 de Febrero arcilla, Santa Ana), pocas tienen obvia preferencia en su distribución justamente a este grupo.
También en el nivel de familias de árboles pueden distinguirse algunas preferencias edáficas. Generalizando un poco, los 15 sectores de estudio pueden arreglarse en una línea que conforma más o menos al orden observado en la ordenación desde Tarapoto hasta Mishana 2 (Figura 7.7A). Si los sectores se organizan en este orden, y se presentan los porcentajes de especies por familia en cada sector (Tabla 7.13), se ve que la distribución de la riqueza relativa de especies por familia no es al azar. Al contrario, hay varias familias cuya diversidad de especies presenta variación según la gradiente edáfica: los valores más altos o más bajos de riqueza de especies se encuentran en una cierta parte de esta gradiente. Entre las 30 familias más ricas en especies, cada una de ellas contiene 10 especies o más (Tabla 7.3), se puede notar que por lo menos 12 familias tienen alguna zona aparentemente preferida en la gradiente edáfica. Flacourtiaceae y Sterculiaceae tienen la mayor concentración de especies en los sectores con suelos arcillosos y con concentraciones de cationes relativamente altas. Burseraceae, Combretaceae, Chrysobalanaceae, Elaeocarpaceae y Lecythidaceae tienen las mayores concentraciones de especies en los lugares con suelos intermedios en términos de textura y concentración de cationes. Cecropiaceae, Meliaceae y Violaceae aparentemente evitan los suelos arenosos y pobres, mientras que Apocynaceae, Myrsinaceae y Sapotaceae justamente son más diversas en tales suelos.
Las abundancias relativas de las familias (Tabla 7.14) proporcionan generalmente las mismas conclusiones de las preferencias edáficas de las familias que la riqueza relativa de especies. Sin embargo, hay algunas familias que presentan tendencias de preferencia en cuanto a su abundancia pero no en cuanto a su riqueza de especies. Por ejemplo las Leguminosae son claramente mas abundantes en los lugares con suelos arenosos y pobres, pero la riqueza de especies no es restringida a suelos pobres. Las Moraseis, en cambio, abundan mas bien afuera de los suelos pobres. Las Melastomatáceas aparecen más abundantes en los lugares con suelos intermedios.
Correlaciones estadísticas entre los patrones florísticos y características del suelo en los 15 sectores de estudio con muestreo de árboles
Las ordenaciones presentadas en la Figura 7.7 dan una idea visual de la correlación entre los patrones florísticos obtenidos a base de los árboles con los patrones obtenidos con los otros grupos de plantas (Melastomatáceas y pteridófitas), y también con los patrones obtenidos con las características de los suelos. Para comprobar esta idea, se han calculado las correlaciones directamente entre las diferentes matrices de similitud. Como se puede ver en la Tabla 7.15, estos análisis dan casi siempre resultados positivos.
Tabla 7.13. Riqueza relativa de especies de 30 familias de árboles en los 15 sectores de estudio. La riqueza relativa está calculada para cada uno de los sectores como porcentaje de especies que pertenecen a la familia en cuestión. Los tres valores más altos para cada familia están en letra negrita. Sólo se incluyen las familias que tenían por lo menos 10 especies de árboles en el presente estudio.
| Familia | Tara- poto | Gen gen | Ex Pet | Mo 2 | San Ant |
13 Feb arci- lla |
Sta Ana | Nauta | Mo 1 | Maniti | Pan gua | Mísh 1 |
13 Feb arena |
Mish 3 | Mish 2 |
| Anacardiaceae |
0,4
|
0,5
|
0,8
|
0,0
|
0,7
|
0,9
|
1,3
|
1,2
|
2,1
|
0,5
|
0,4
|
1,0
|
1,5
|
1,5
|
1,3
|
| Annonaceae |
7,5
|
6,0
|
8,3
|
6,7
|
4,8
|
6,1
|
4,7
|
5,7
|
4,8
|
5,1
|
4,6
|
5,6
|
6,8
|
6,8
|
5,0
|
| Apocynaceae |
1,3
|
1,4
|
2,5
|
1,2
|
1,8
|
1,8
|
0,9
|
1,6
|
1,4
|
1,9
|
2,5
|
2,6
|
3,0
|
3,0
|
2,5
|
| Acercare |
2,2
|
1,8
|
2,1
|
1,8
|
1,1
|
2,6
|
2,6
|
3,3
|
1,4
|
2,8
|
2,1
|
1,5
|
2,3
|
3,0
|
0,0
|
| Bombacaceae |
1,3
|
2,3
|
2,1
|
0,6
|
1,1
|
1,8
|
1,7
|
1,6
|
0,0
|
1,4
|
0,4
|
0,0
|
1,5
|
0,8
|
0,6
|
| Burseraceae |
4,4
|
2,3
|
3,3
|
7,9
|
4,4
|
5,3
|
4,7
|
4,9
|
9,0
|
1,9
|
6,7
|
7,2
|
7,5
|
3,0
|
2,5
|
| Cecropiaceae |
2,2
|
1,8
|
1,7
|
1,8
|
2,6
|
1,8
|
1,3
|
4,1
|
2,1
|
3,3
|
2,1
|
2,1
|
0,8
|
0,8
|
0,6
|
| Chrysobalanaceae |
0,0
|
2,8
|
1,7
|
1,2
|
3,0
|
2,6
|
6,0
|
4,5
|
2,1
|
4,7
|
4,6
|
4,1
|
3,8
|
2,3
|
2,5
|
| Clusiaceae |
3,5
|
1,4
|
0,8
|
1,2
|
2,2
|
0,9
|
0,4
|
3,3
|
2,8
|
3,7
|
2,1
|
3,6
|
0,0
|
3,0
|
3,1
|
| Combretaceac |
0,0
|
0,0
|
0,4
|
0,6
|
0,0
|
0,9
|
0,0
|
0,4
|
2,1
|
0,5
|
1,7
|
0,5
|
0,8
|
1,5
|
1,3
|
| Elaeocarpaceae |
0,4
|
1,4
|
1,3
|
0,6
|
1,1
|
0,0
|
2,2
|
2,9
|
6,9
|
2,3
|
3,3
|
2,1
|
5,3
|
3,0
|
0,6
|
| Euphorbiaceae |
3,1
|
3,2
|
3,8
|
3,6
|
4,4
|
3,5
|
3,9
|
3,3
|
2,8
|
3,3
|
0,8
|
2,1
|
8,3
|
3,8
|
3,8
|
| Flacourtiaceae |
2,6
|
4,1
|
3,3
|
3,6
|
2,6
|
2,6
|
2,2
|
2,4
|
1,4
|
1,4
|
0,8
|
0,0
|
1,5
|
1,5
|
1,9
|
| Lauraceae |
5,7
|
5,5
|
3,3
|
7,9
|
4,1
|
3,5
|
6,5
|
3,3
|
8,3
|
5,6
|
7,9
|
8,7
|
1,5
|
9,1
|
7,5
|
| Lecythidaceae |
2,2
|
1,8
|
1,7
|
3,6
|
3,3
|
2,6
|
4,7
|
4,9
|
3,4
|
2,8
|
2,9
|
3,1
|
2,3
|
0,0
|
0,0
|
| Leguminosae |
9,7
|
8,3
|
12,5
|
9,1
|
11,1
|
7,0
|
10,3
|
10,6
|
4,8
|
9,8
|
10,5
|
10,3
|
15,0
|
9,1
|
14,5
|
| Melastomataceae |
1,3
|
2,3
|
1,3
|
1,2
|
0,4
|
4,4
|
3,0
|
2,4
|
2,1
|
1,9
|
5,4
|
2,6
|
1,5
|
1,5
|
0,6
|
| Meliaecae |
5,7
|
6,4
|
6,3
|
4,8
|
4,8
|
3,5
|
3,0
|
2,0
|
3,4
|
4,2
|
5,0
|
5,6
|
1,5
|
0,8
|
1,3
|
| Monimiaceae |
0,4
|
1,4
|
1,7
|
3,0
|
2,2
|
2,6
|
1,3
|
1,2
|
0,7
|
1,9
|
0,4
|
1,0
|
0,8
|
1,5
|
0,0
|
| Moraceae |
5,7
|
6,0
|
5,4
|
4,2
|
7,0
|
5,3
|
2,2
|
5,7
|
6,2
|
5,6
|
2,9
|
3,6
|
4,5
|
5,3
|
0,6
|
| Myristicaceae |
5,7
|
5,5
|
4,6
|
4,8
|
5,2
|
10,5
|
3,9
|
3,7
|
8,3
|
7,5
|
4,6
|
7,7
|
6,0
|
5,3
|
2,5
|
| Myrsinaceae |
0,4
|
0,5
|
0,4
|
0,0
|
0,0
|
0,0
|
0,0
|
0,0
|
0,0
|
0,0
|
0,8
|
1,0
|
0,8
|
0,8
|
2,5
|
| Myrtaceae |
2,6
|
4,6
|
4,6
|
3,0
|
2,6
|
0,9
|
4,7
|
2,0
|
1,4
|
3,3
|
2,1
|
1,5
|
1,5
|
4,5
|
6,3
|
| Nyctaginaceae |
1,8
|
1,4
|
2,5
|
1,8
|
1,5
|
2,6
|
1,3
|
0,8
|
0,0
|
1,4
|
1,3
|
0,0
|
0,8
|
1,5
|
2,5
|
| Olacaceae |
0,9
|
0,9
|
0,8
|
2,4
|
1,1
|
1,8
|
0,0
|
1,2
|
0,7
|
0,9
|
0,8
|
1,5
|
0,8
|
0,8
|
1,3
|
| Rubiaceae |
4,8
|
5,0
|
5,0
|
5,5
|
2,6
|
4,4
|
4,3
|
4,5
|
2,1
|
2,8
|
5,0
|
4,1
|
2,3
|
5,3
|
3,8
|
| Sapindaecae |
2,6
|
0,5
|
1,7
|
0,0
|
2,2
|
0,0
|
0,4
|
0,8
|
0,7
|
0,9
|
0,8
|
0,5
|
1,5
|
2,3
|
1,9
|
| Sapotaceae |
6,6
|
5,0
|
3,8
|
6,1
|
7,7
|
7,9
|
7,8
|
8,6
|
4,8
|
6,5
|
4,6
|
7,2
|
6,0
|
6,1
|
14,5
|
| Sterculiaceae |
1,8
|
1,8
|
2,1
|
1,8
|
2,6
|
1,8
|
1,3
|
0,4
|
1,4
|
0,9
|
0,8
|
1,5
|
1,5
|
0,0
|
0,0
|
| Violaceae |
2,2
|
1,4
|
1,3
|
2,4
|
2,6
|
1,8
|
2,2
|
1,6
|
1,4
|
2,3
|
1,3
|
1,0
|
0,0
|
0,0
|
0,0
|
Especialmente, las tres matrices florísticas están fuertemente correlacionadas entre si. La matriz general del suelo (calculada en base a todas las características medidas en los 15 sectores de muestreo) está también claramente correlacionada con las matrices florísticas. De las diferentes características del suelo, son el porcentaje de arena y el contenido de potasio los que correlacionan con los tres grupos florísticos. Además, LOI y aluminio tienen correlaciones elevadas con árboles y pteridófitas. Los cationes extraibles están correlacionadas más que todo con árboles y Melastomatáceas. La distancia geográfica no tuvo mucho que hacer con la composición florística. Aunque tiene una correlación significativa con las pteridófitas,
esta correlación es negativa y antes que todo es resultado de que algunos de los sectores de muestreo, que tenían características de suelo y flora muy disimilares (por ejemplo 13 de Febrero arena y arcilla), quedaban geográficamente muy cerca entre sí.
Tabla 7.14. Abundancia relativa de 30 familias de árboles en los 15 sectores en estudio. La abundancia relativa está calculada para cada uno de los sectores como porcentaje de individuos que pertenecen a la familia en cuestión. Para otros detalles, véase Tabla 7.15.
| Familia | Tara- poto | Gen gen | Ex Pet | Mo 2 | San Ant |
13 Feb arci- lla |
Sta Ana | Nauta | Mo 1 | Mani- ti | Pan gua | Mís h 1 |
13 Feb arena |
Mis 3 | Mis 2 |
| Anacardiaecae |
0,2
|
0,2
|
1,0
|
0,0
|
0,4
|
1,3
|
0,8
|
0,9
|
1,3
|
0,2
|
0,2
|
0,6
|
1,5
|
0,5
|
0,2
|
| Annonaceae |
4,7
|
3,5
|
9,3
|
5,2
|
2,6
|
5,3
|
3,8
|
4,9
|
6,7
|
4,8
|
5,3
|
3,8
|
7,0
|
4,4
|
4,9
|
| Apocynaceae |
1,0
|
0,9
|
2,0
|
0,9
|
1,1
|
2,0
|
0,8
|
1,1
|
0,7
|
0,9
|
1,8
|
0,9
|
2,5
|
5,5
|
4,7
|
| Acercare |
3,5
|
2,8
|
2,3
|
1,2
|
0,9
|
2,7
|
2,7
|
3,4
|
1,3
|
5,4
|
7,3
|
3,8
|
2,5
|
3,4
|
0,0
|
| Bombacaecae |
1,0
|
1,6
|
1,5
|
0,6
|
1,1
|
2,0
|
1,1
|
1,1
|
0,0
|
4,6
|
0,9
|
0,0
|
1,5
|
1,6
|
3,0
|
| Burseraceae |
3,7
|
3,9
|
5,0
|
7,3
|
4,2
|
8,7
|
4,1
|
5,8
|
10,8
|
1,7
|
6,4
|
6,0
|
10,4
|
2,3
|
0,6
|
| Ceeropiaceae |
1,2
|
1,6
|
1,0
|
1,2
|
1,5
|
1,3
|
1,1
|
3,4
|
1,0
|
2,6
|
1,4
|
2,3
|
1,0
|
0,2
|
0,4
|
| Chrysobalanaceae |
0,0
|
2,8
|
2,5
|
1,8
|
2,6
|
2,0
|
5,2
|
3,6
|
1,7
|
4,6
|
4,4
|
3,0
|
4,5
|
7,1
|
1,6
|
| Clusiaceae |
2,2
|
0,7
|
0,8
|
1,2
|
1,5
|
0,7
|
0,3
|
2,5
|
1,3
|
2,0
|
1,8
|
3,6
|
0,0
|
1,8
|
3,4
|
| Combretaceac |
0,0
|
0,0
|
0,3
|
0,3
|
0,0
|
0,7
|
0,0
|
0,2
|
1,0
|
0,2
|
1,4
|
0,2
|
0,5
|
0,5
|
0,2
|
| Elaeocarpaceae |
0,2
|
0,7
|
0,8
|
0,6
|
0,6
|
0,0
|
1,4
|
2,2
|
4,4
|
1,1
|
2,8
|
1,3
|
4,0
|
1,1
|
0,1
|
| Euphorbiaceae |
3,5
|
3,5
|
5,5
|
3,6
|
10,0
|
3,3
|
10,1
|
3,8
|
4,7
|
10,9
|
2,1
|
6,2
|
11,9
|
2,5
|
6,9
|
| Flacourtiaceae |
1,5
|
4,4
|
2,5
|
8,5
|
3,0
|
3,3
|
1,6
|
2,2
|
0,7
|
1,3
|
0,5
|
0,0
|
1,0
|
1,1
|
1,9
|
| Lauraceae |
3,2
|
4,2
|
2,8
|
6,1
|
2,4
|
2,7
|
4,6
|
1,8
|
7,4
|
3,9
|
6,0
|
5,1
|
1,0
|
4,4
|
2,4
|
| Lecythidaceae |
3,0
|
4,4
|
1,8
|
2,1
|
4,2
|
4,0
|
4,9
|
6,7
|
6,1
|
5,0
|
6,0
|
4,1
|
2,0
|
0,0
|
0,0
|
| Leguminosae |
9,4
|
6,9
|
11,3
|
8,2
|
9,0
|
5,3
|
9,0
|
12,8
|
3,4
|
8,1
|
7,8
|
7,7
|
13,9
|
17,5
|
13,6
|
| Melastomataceae |
0,7
|
1,6
|
1,0
|
1,8
|
0,2
|
3,3
|
3,3
|
2,0
|
3,0
|
1,7
|
4,8
|
4,1
|
1,5
|
0,9
|
0,1
|
| Meliaceae |
7,7
|
8,8
|
6,3
|
4,2
|
3,0
|
4,0
|
2,7
|
1,6
|
2,0
|
3,9
|
4,6
|
7,7
|
1,0
|
0,7
|
1,0
|
| Monimiaceae |
0,5
|
3,5
|
1,0
|
3,9
|
2,8
|
4,7
|
1,4
|
2,7
|
2,4
|
1,3
|
0,5
|
0,4
|
0,5
|
0,7
|
0,0
|
| Moraceae |
4,5
|
3,9
|
5,8
|
3,0
|
6,6
|
4,0
|
1,9
|
5,8
|
4,4
|
3,7
|
2,3
|
3,0
|
3,5
|
2,1
|
0,2
|
| Myristicaceae |
9,7
|
9,9
|
5,3
|
10,3
|
7,0
|
10,7
|
4,9
|
7,8
|
13,8
|
8,3
|
8,3
|
13,0
|
6,5
|
5,1
|
3,4
|
| Myrsinaceae |
0,2
|
0,2
|
0,3
|
0,0
|
0,0
|
0,0
|
0,0
|
0,0
|
0,0
|
0,0
|
0,7
|
0,4
|
0,5
|
0,2
|
1,4
|
| Myrtaceae |
2,2
|
2,8
|
3,0
|
2,1
|
1,7
|
0,7
|
4,4
|
1,8
|
1,0
|
1,7
|
1,6
|
0,6
|
1,0
|
3,0
|
3,5
|
| Nyctaginaceac |
1,0
|
1,8
|
2,3
|
3,3
|
1,3
|
2,7
|
1,1
|
0,7
|
0,0
|
2,2
|
1,4
|
0,0
|
0,5
|
1,1
|
5,9
|
| Olacaceae |
1,0
|
1,6
|
1,3
|
1,5
|
1,8
|
2,0
|
0,0
|
1,1
|
0,3
|
0,9
|
0,9
|
1,3
|
1,0
|
0,2
|
0,4
|
| Rubiaceae |
6,2
|
6,2
|
5,3
|
4,8
|
2,4
|
3,3
|
3,3
|
2,9
|
6,1
|
1,7
|
4,8
|
1,9
|
2,5
|
3,0
|
3,5
|
| Sapindaecae |
2,0
|
0,2
|
1,8
|
0,0
|
1,3
|
0,0
|
0,3
|
1,1
|
0,3
|
0,4
|
0,7
|
0,6
|
1,5
|
1,8
|
2,9
|
| Sapotaceae |
5,2
|
3,0
|
3,0
|
3,6
|
4,8
|
6,7
|
6,3
|
5,6
|
3,4
|
4,4
|
4,4
|
5,1
|
4,0
|
12,2
|
14,5
|
| Sterculiaceae |
1,7
|
2,5
|
1,5
|
1,2
|
1,7
|
1,3
|
1,4
|
0,2
|
1,0
|
1,1
|
1,1
|
1,3
|
2,0
|
0,0
|
0,0
|
| Violaceae |
9,7
|
1,8
|
2,5
|
2,4
|
11,1
|
2,7
|
4,9
|
4,0
|
3,0
|
4,1
|
0,9
|
5,3
|
0,0
|
0,0
|
0,0
|
Tabla 7.15. Correlaciones entre diferentes matrices de similitud según la prueba de Mantel. Los análisis se han hecho utilizando los 15 sectores de estudio donde se hicieron inventarios de árboles. En el caso de fósforo, las correlaciones están basadas en 7 (P-Bray) ú 8 (P) sectores, porque los dos diferentes métodos utilizados para medir el fósforo no dan resultados comparables. Para obtener las probabilidades de error (p), se han utilizado 999 permutaciones: * p <0,05; ** p <0,01; *** p <0,001.
| Árboles | Melastomatáceas | Pteridófitas | |
| Melastomatáceas |
0,61***
|
|
|
| pteridófitas |
0,75***
|
0,65***
|
|
| Ca, 1,, Nig, Na, Al, pH, LOI, arena |
0,56***
|
0,32**
|
0,57***
|
| Arena |
0,38**
|
0,19*
|
0,44**
|
| Ca, K, Mg, Na |
0,42***
|
0,28*
|
0,29
|
| Ca |
0,31*
|
0,21
|
0,15
|
| K |
0,47***
|
0,31**
|
0,36**
|
| Nig |
0,31*
|
0,22
|
0,16
|
| Na |
0,23*
|
0,21
|
0,24
|
| Al |
0,36**
|
0,07
|
0,47**
|
| pH |
0,21*
|
0,18
|
0,30
|
| LOI |
0,47***
|
0,17
|
0,54***
|
| Distancia geográfica |
-0,13
|
-0,00
|
-0,30***
|
| P-Bray |
0,24
|
0,49*
|
0,18
|
| p |
0,19
|
0,05
|
0,63*
|
Ordenaciones y caracterizaciones de los 32 sectores de estudio con inventarios de Melastomatáceas y pteridófitas
Las Melastomatáceas y pteridófitas se estudiaron no sólo en los 15 sectores de muestreo donde se hicieron los inventarios de árboles, sino también en 17 sectores adicionales. Las tres ordenaciones de estos 32 lugares, obtenidas en base a Melastomatáceas, pteridófitas y características del suelo, se presentan en la Figura 7.8. En cada una de las tres ordenaciones se pueden dividir los sectores de muestreo en seis grupos de una manera casi igual. Especialmente las ordenaciones florísticas producen imágenes muy semejantes: en el primer grupo, al extremo izquierdo, se encuentran Palo Seco 1, 7 de Julio, Tarapoto, Gengen, Libertad Agraria km8 y Ex Petroleros. Carbajal km0 y km2 juntos con San Antonio 1 forman el segundo grupo, y cerca de este grupo hay cuatro lugares más: Palo Seco 2, Momón 2, 13 de Febrero arcilla y Santa Ana. El cuarto grupo se sitúa, en ambas ordenaciones, en la parte central superior de la imagen y contiene seis lugares: Carbajal km4, km8, km. 10 y km. 12 más Nauta y San Antonio 2.
Las mayores diferencias entre las ordenaciones florísticas se muestran en la parte derecha de la ordenación. En base a las pteridófitas, se pueden distinguir claramente dos grupos. Más cerca de los grupos anteriores están Maniti, Panguana 1 y 2, Libertad Agraria km3, Momón 1, Mishana 1 y San Pedro. El último grupo tiene seis lugares: 13 de Febrero arena, San Gerardo, Carbajal km. 6, Puerto Almendras, Mishana 2 y Mishana 3. En base a las Melastomatáceas se ve una constelación algo semejante, pero las posiciones de Carbajal km6, Momón 1 y Mishana 3, difieren claramente del patrón de las pteridófitas.
En la ordenación basada en las características del suelo, se ven muchos de los mismos grupos de lugares que ya fueron indicados en las ordenaciones florísticas. Sin embargo, las posiciones relativas de los grupos son un poco diferentes y cuatro lugares (Gengen, Momón 2, Nauta y Palo Seco 2) se encuentran claramente en posiciones diferentes a las que dan las ordenaciones florísticas. Igual como en el caso de los 15 lugares con árboles, también aquí la secuencia de lugares de izquierda a derecha corresponde a disminución de la concentración de cationes y arcilla en el suelo (Tabla 7.11).
Entre las Melastomatáceas que se han podido identificar hasta la especie, hay unas 14 especies comunes, las cuales han sido observadas en varios sectores de estudio, pero solamente dentro de uno de los seis grupos de sectores de muestreo definidos anteriormente (Tabla 7.16, Figura 7.8). Como cada grupo tiene características edáficas o geológicas distintas, estas especies con distribución restringida, pueden considerarse como especialistas, y pueden ser utilizadas como indicadoras para ciertas condiciones ecológicas (véase Capítulo 13). En los grupos 1, 4 y 5 ocurren varias especies que son características para un solo grupo y no han sido observadas dentro de los otros grupos. Se nota que no hay especies específicas para los grupos 2 o 3. Algunas especies se han encontrado en todos los grupos desde 1 hasta 3 6 4 (Clidemia epiphytica, Miconia grandillona, Miconia serrulata y Ossaea boliviensis). La única especie que se encuentra frecuentemente en todos los grupos desde el 2 hasta el 6, pero no en el grupo 1, es Miconia klugii. Las otras especies que se encuentran en el rango de grupos desde 2 hasta 5 o 6, también se encuentran en el grupo 1 (por ejemplo Maieta guianensis, Miconia prasina, Miconia tomentosa y Tococa ulei). Eso significa que estas especies son generalistas que abundan practicablemente en todas partes, y por eso sus distribuciones no tienen valor indicativo en cuanto a las condiciones edáficas.
Hay tres especies de Miconia que se han encontrado exclusivamente en los grupos 1 y 3. En la mayoría de los casos, especies compartidas entre estos grupos también ocurren en el grupo 2. Aunque los grupos 2 y 3 comparten varias especies, en términos generales se puede decir que las especies que ocurren en el grupo 2 tienen una tendencia de crecer también en el grupo 1, mientras que las especies comunes en el grupo 3 comúnmente no ocurren en, grupos 1 y 2 pero sí en los grupos 4-6 (por ejemplo, Miconia dolichorrhyncha, Miconia rimachii, Miconia tetrasperma, Miconia traillii y Tococa guianensis).
Clidemia epibaterium es la única especie que claramente prefiere los sectores del grupo 6 (Tabla 7.16), pero hay varias especies que crecen en sectores de los grupos 6 y 5 (Clidemia longifolia, Miconia barbinervis y Ossaea cucullata) o además en el grupo 4 (Adelobotrys marginata, Miconia carassana, Miconia pterocaulon y Ossaea araneifera).
En las pteridófitas hay unas 23 especies comunes que han sido observadas en más de un sector de estudio, pero solamente dentro de uno de los seis grupos de sectores de muestreo definidos arriba (Tabla 7.17, Figura 7.8). Como cada grupo tiene características edáficas o geológicas distintas, estas especies con distribución restringida pueden ser consideradas como especialistas, y pueden ser utilizadas como indicadoras para ciertas condiciones ecológicas (véase Capítulo 13). Espe285
cialmente los grupos 1 y 6 tienen varias especies especialistas que no han sido observadas dentro de lugares de los otros grupos. Se nota que no hay especies específicas para los grupos 2 6 3. Hay varias especies restringidas a dos grupos de lugares, especialmente los que ocurren tanto en el grupo 1 como en el 2, y otros que ocurren en los grupos 5 y 6 (Tabla 7.17).
Tabla 7.16. Especies de Melastomataceae que se han identificadas hasta la especie y que tienen una clara preferencia a uno o dos grupos de sectores de estudio. Los seis círculos representan, de izquierda a derecha, los grupos 1 a 6 (véase Figura 7.8), y más o menos corresponden a la gradiente edáfica desde suelos arcillosos con contenido elevado de cationes a la izquierda hacia suelos arenosos con contenido bajo de cationes a la derecha. Círculos negros significan que las especies mencionadas al costado han sido observadas en la parte respectiva de la gradiente, círculos blancos significan lo contrario.
| Grupos preferidos | Especies |
| ©OOOOO | Adelobotrys macrantha, Clidemia heterophylla, Clidemiaseptuplinervia, Miconia napoana, Triolena amazonica |
| ©©OOOO | Clidemia dimorphica, Miconia aureoides, Leandra longicoma,Tococa caquetana |
| ©O©OOO | Miconia elata, Miconia lamprophylla, Miconia trinervia |
| O©OOOO | .- |
| O©O©OO | Miconia dispar, Adelobotrys macrophylla, micronia umbriensis |
| OO©OOO | .- |
| OO©©OO | Miconia ampla |
| OO©O©O | Bellucia grossularioides, Clidemia piperifolia, Leandra chaetodon, Miconia crassinervia |
| OOO©OO | Miconia centrodesma, Miconia egensis, Miconia schunkei |
| OOO©©O | Miconia duckei, Miconia phanerostila, Miconia rugosa, Miconia tetragona |
| OOOO©O | Adelobotrys praetexta, Leandra nanayensis, Miconia lepidota,Miconia spichigeri, Tococa caryophyllea |
| OOOO©© | Miconia myriantha, Miconia barbinervis |
| OOOOO© | Clidemia epibaterium |
En los casos cuando una especie ocurre en tres grupos de lugares, el patrón más común es que ocurren en los grupos 1-3 (siete especies, por ejemplo Polybotrya crassirhizoma, Saccoloma elegans y Microgramma fuscopunctata). Cuatro especies son restringidas a los grupos 4-6 (incluso Adiantum tomentosum, Lindsaea falcata, Cyathea bradei), y tres especies a cada uno de las siguientes combinaciones de grupos: 1,2, y 4; 2, 3 y 4; 3, 5 y 6. Dos especies son restringidas a los grupos 3, 4 y 5, y cuatro especies se han encontrado en alguna otra combinación de tres grupos. De estos patrones se puede apreciar claramente que la mayoría de las especies tiene un patrón de distribución que parece ecológicamente determinado. El contenido promedio de nutrientes en el suelo disminuye del grupo 1 hacia grupo 6, y la mayoría de las especies que ocurren en más de un grupo, han sido encontradas en grupos consecutivos, o sea, en grupos que tienen los suelos más parecidos. No hay ni una especie que ocurre en los dos grupos extremos en cuanto a esta variación edáfica sin ocurrir en todos o casi todos los grupos intermedios.
 |
Figura 7.8. Ordenaciones de coordenadas principales de los 32 sectores de estudio basados en características diferentes. A. composición florística de Melastomatáceas (índice de Jaccard). B. composición florística de pteridófitas (índice de Jaccard). C. características del suelo (distancia euclidiana utilizando pH, porcentaje de arena, y el contenido de Al, Ca, K, Mg y Na). Los seis grupos de sectores (numerados del 1 al 6) que más o menos reaparecen en todas las ordenaciones han sido separados con criterios visuales. |
Hay nueve especies que pueden ser consideradas muy generalistas, ya que ocurren en todos los seis grupos de lugares de estudio (por ejemplo, Cyathea lasiosora, Nephrolepis rivularis, Lomariopsis nigropaleata, Metaxya rostrata, Cyclodium meniscioides). Además hay 11 especies generalistas que ocurren en todos los grupos de lugares excepto en el grupo.6 (por ejemplo, Adiantum terminatum, Cyathea pungens, Lomagramma guianensis, Polybotrya osmundacea, Salpichlaena volubilis), y otras cuatro especies que ocurren en todos los grupos excepto el 1 (Polybotrya pubens, Scizaea elegans, Selaginella parkeri, Lindsaea lancea).
Tabla 7.17. Ejemplos de especies de pteridófitas que solamente han sido observadas dentro de uno o dos grupos de sectores de estudio. Los seis círculos representan, de izquierda a derecha, los grupos 1-6 (véase Figura 7.8), y más o menos corresponden a la gradiente edáfica desde suelos arcillosos con contenido de cationes elevado a la izquierda hacia suelos arenosos con contenido de cationes bajo a la derecha. Los círculos negros significan que las especies mencionadas al costado han sido observadas en la parte respectiva de la gradiente, círculos blancos significan lo contrario.
| Grupospreferidos | Especies |
| ©OOOOO | Bolbitis lindigii, Cyathea amazonica, Thelypteris biformata,Thelypteris pennata, Diplazium striatum, Lomariopsis fendleri,Pteris pungens, Cyclodium trianae, Didymochlaena truncatula |
| ©©OOOO | Adiantum pulverulentum, Cyathea cuspidata, Selaginellahaematodes, Tectaria antioquoiana, Thelypteris tristis |
| ©O©OOO | Polypodium decumanum, Pteris propinqua |
| O©OOOO | -- |
| O©©OOO | Trichomanes hostmannianum |
| O©O©OO | Lomariopsis latipinna |
| OO©OOO | -- |
| OO©©OO | Lindsaea taeniata |
| OO©O©O | Selaginella conduplicata |
| OOO©OO | Cnemidaria ewanii |
| OOOO©O | Salpichlaena hookeriana, Selaginella palmiformis |
| OOOO©© | Asplenium hallii, Elaphoglossum styriacum, Lindsaea hemiglossa, Trichomanes accedens |
| OOOOO© | Trichomanes martiusii, Trichomanes bicorne, Arachniodes macrostegia, Lindsaea tetraptera |
Correlaciones entre patrones florísticos y características del suelo
Las ordenaciones ya han sugerido que en los 32 sectores de estudio, la correlación entre las Melastomatáceas y las pteridófitas es más fuerte que las correlaciones entre la flora y las características de suelo. Loó resultados de los análisis de correlación entre las diferentes matrices de similitud confirman esta observación (Tabla 7.18). En general, las pteridófitas corresponden mejor a la variación en el suelo que las Melastomatáceas, pero la diferencia no es tan marcada como cuando sólo se tomaban en cuenta los 15 sectores de muestreo donde también se habían muestreado árboles (Tabla 7.15). La variación florística, tanto en pteridófitas como en Melastomatáceas, generalmente corresponde a las mismas características del suelo. Variación en la textura (el contenido de arena) y en el contenido de calcio, magnesio y potasio son las características más claramente relacionadas con la variación florística. Fósforo y aluminio corresponden un poco menos a los cambios florísticos, y sodio, pH y LOI tienen muy poca correlación con la flora.
Tabla 7.18. Correlaciones entre las diferentes matrices de similitud según la prueba de Mantel. La mayoría de los análisis se han hecho utilizando todos los 32 sectores en estudio donde hay inventarios de Melastomatáceas y pteridófitas (compare con los resultados presentados en Tabla 7.15, donde solamente se han tomado en cuenta los 15 sectores con muestreo de árboles). En el caso del fósforo (P-Bray), las correlaciones están basadas en 24 sectores, y en el caso de LOI en 31 sectores, porque no se han analizado estas características en los 32 sectores. Probabilidades de error, ver la Tabla 7.15.
| . | Melastomatáceas | Pteridófitas |
| Pteridófitas |
0,65***
|
|
| Ca, K, Mg, Na, Al, pH, arena |
0,32***
|
0,45***
|
| Arena |
0,26***
|
0,36***
|
| Ca, K, Mg, Na |
0,28***
|
0,40***
|
| Ca |
0,33***
|
0,45***
|
| K |
0,30***
|
0,36***
|
| Mg |
0,34***
|
0,38***
|
| Na |
-0,01
|
0,05
|
| Al |
0,11*
|
0,26**
|
| pH |
0,12
|
0,16
|
| Distancia geográfica |
0,16**
|
0,04
|
| LOI |
0,04
|
0,16
|
| P-Bray |
0,22**
|
0,20**
|
CORRELACIONES CON LOS PATRONES EN LA IMAGEN DE SATÉLITE LANDSAT TM
Arboles
Se podía registrar información digital de reflectancias de luz en 10 de los 15 sectores donde se han hecho inventarios de árboles. En cinco sectores este registro no fue posible debido a la deforestación (Momón 1 y 2), por dificultades en determinar la posición geográfica del lugar (Mishana 3) ó por cobertura de nubes (Panguana 1 y Tarapoto).
Las ordenaciones de los diez lugares basados en árboles y en base a la combinación de las bandas 1-5 y 7 en la imagen de satélite Landsat TM resultan en un patrón muy semejante (Figura 7.9). Prácticamente, es sólo la posición de Mishana 1 la que significativamente difiere entre estas dos ordenaciones. La correlación calculada entre árboles y la misma combinación de bandas da también un valor relativamente alto (Tabla 7.19). Considerando cada una de las bandas separadamente, se ve que las bandas que representan luz infrarroja (bandas 4, 5 y 7), se correlacionan mucho mejor con la variación florística que en las bandas de luz visible (bandas 1, 2 y 3). Consecuentemente, la combinación de las tres bandas de luz infrarroja, resulta en una mejor correlación con la flora que la correlación obtenida en base a la combinación de las bandas de luz visible.
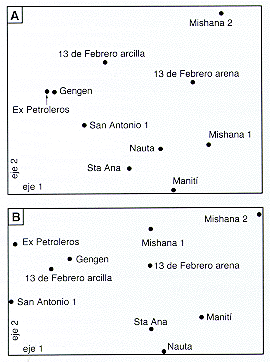 |
Figura 7.9. Las ordenaciones de coordenadas principales de los 10 sectores con muestreo de árboles donde también se han podido registrar las reflectancias de luz en una imagen de satélite Landsat TM. Ordenaciones basadas en composición florística de árboles (índice de Jaccard). B. combinación de las bandas 1-5 y 7 de Landsat TM (distancia euclidiana). |
Tabla 7.19. Correlaciones entre diferentes matrices de similitud según la prueba de Mantel. Las matrices de similitud han sido calculadas basadas en las reflectancias de luz en diferentes bandas de una imagen de satélite Landsat TM, y en base a la composición florística de árboles en los 10 sectores en estudio. Probabilidades de error, ver la Tabla 7.15.
| BANDAS DEL LANDSAT TM | Árboles |
| Bandas 1-5, 7 | 0,57** |
| Bandas 1,2,3 | 0,22 |
| Bandas 4,5,7 | 0,55** |
| Banda 1 | 0,10 |
| Banda 2 | 0,28 |
| Banda 3 | 0,21 |
| Banda 4 | 0,43** |
| Banda 5 | 0,59** |
| Banda 7 | 0,44** |
Melastomatáceas y pteridófitas
Se podía registrar información digital de reflectancias de luz en 23 de los 32 sectores donde se han hecho los inventarios de Melastomatáceas y pteridófitas. En nueve sectores este registro no fue posible por cobertura de nubes, problemas en la localización geográfica del sector estudiado, o por deforestación.
Las ordenaciones basadas en la composición florística de Melastomatáceas (Figura 7.10A) y pteridófitas (Figura 7.10B) son muy semejantes a las ordenaciones en las Figuras 7.8A y 7.8B, respectivamente, y se pueden distinguir los mismos grupos de sectores de estudio. Esto es lógico ya que los datos utilizados para conseguir las ordenaciones son casi los mismos: la diferencia es que en la Figura 7.8 se han utilizado los 32 sectores de muestreo, mientras que en la Figura 7.10 fue necesario omitir los 9 sectores de muestreo donde no fue posible obtener valores de reflectancia en la imagen de satélite. La gradiente edáfica corresponde más o menos al orden de los grupos de la izquierda, donde se encuentran los sectores con suelos arcillosos (por ejemplo Gengen y Ex Petroleros), hacía los grupos de sectores con suelos intermedios (por ejemplo San Antonio 2 y Nauta) hasta el grupo de sectores con suelos arenosos (Mishana 2). Sólo en el caso de las Melastomatáceas, tres sectores de muestreo (Santa Ana, 13 de Febrero arcilla y Carbajal km6) adoptan posiciones que no corresponden muy bien a esta gradiente.
En la ordenación de los estos 23 sectores en estudio en base a las reflectancias de luz en la imagen de satélite (Figura 7.10C), se puede reconocer en términos generales el mismo arreglo, aunque hay excepciones notables. Las discrepancias más obvias son ,as posiciones de 7 de julio y Libertad Agraria km8. Según sus características florísticas, estos dos sectores pertenecen a la parte extrema izquierda de la ordenación juntos con Ex Petroleros y Gengen, pero en base a las reflectancias resultan relativamente intermedios y semejantes a por ejemplo Nauta, Santa Ana y Carbajal km6.
 |
Figura 7.10. Ordenaciones de coordenadas principales basadas en diferentes características de los 23 sectores con muestreo de Melastomatáceas y Pteridófitas donde también se han podido registrar las reflectancias de luz en una imagen de satélite Landsat TM. A. Composición florística de Melastomatáceas (índice de Jaccard). B. Composición florística de Pteridófitas (índice de Jaccard). C. Combinación de las bandas 1-5 y 7 de Landsat TM (distancia euclidiana). |
Este resultado se debe a que la parte oriental de la imagen Landsat TM tiene una tonalidad generalmente más oscura que la parte occidental, y se puede observar que todos los sectores en estudio que están geográficamente situados al este del río Amazonas se dibujan en la Figura 7.10C en el extremo superior derecho. Si se comparan sólo estos sectores de estudio entre si, el patrón obtenido por las reflectancias corresponde más o menos al patrón florístico, aunque la posición de Libertad Agraria km8 es diferente. También, si se comparan entre si sólo los sectores de estudio situados al lado occidental del río Amazonas, las ordenaciones florísticas y de reflectancia de luz si corresponden. La división de la región por el río Amazonas en la imagen de satélite es algo permanente, ya que es visible en imágenes tomadas en diferentes años y por diferentes satélites. Sin embargo, queda sin explicación, porque no hay diferencias florísticas o edáficas aparentes que correspondan a esta diferencia en reflectancia.
Las correlaciones calculadas entre las matrices de similitud basadas en la composición florística de Melastomatáceas o Pteridófitas y a las reflectancias de luz, resultan casi siempre en correlaciones positivas y estadísticamente significativas (Tabla 7.20). En el caso de las Melastomatáceas, los valores de correlación son generalmente mayores que los valores obtenidos con Pteridófitas, pero especialmente en las bandas de luz infrarroja no llegan a ser tan altos como los valores obtenidos en el caso de los árboles (Tabla 7.19). En general, los árboles presentan las correlaciones más altas con las bandas de luz infrarroja, mientras que las Melastomatáceas pteridófitas están más correlacionadas con las bandas de luz visible.
Tabla 7.20. Correlaciones entre diferentes matrices de similitud según la prueba de Mantel. Las matrices de similitud han sido calculadas basadas en la reflectancias de luz en diferentes bandas de una imagen de satélite Landsat TM, y en base a la composición florística de Melastomatáceas o Pteridófitas en los 23 sectores de estudio. Probabilidades de error, ver la Tabla 7.15.
|
BANDAS DE LANDSAT TM, UTILIZADAS PARA CALCULAR LA MATRIZ |
Melastomatáceas
|
Pteridófitas
|
| Bandas 1-5, 7 | 0,42*** | 0,32** |
| Bandas 1,2,3 | 0,40*** | 0,36* |
| Bandas 4,5,7 | 0,31*** | 0,21* |
| Banda 1 | 0,35*** | 0,29* |
| Banda 2 | 0,38*** | 0,37* |
| Banda 3 | 0,31** | 0,29* |
| Banda 4 | 0,29** | 0,21** |
| Banda 5 | 0,29*** | 0,12* |
| Banda7 | 0,23** | 0,07 |
RIQUEZA DE ESPECIES
El número de especies de árboles encontrado en este estudio (1416 especies en 6253 individuos), presenta un valor muy alto en comparación con otros registros sobre la riqueza local o regional de especies vegetales en las selvas bajas neotropicales. La diversidad observada es comparable con los resultados de inventarios locales de árboles en parcelas de 1 ó 0,1 hectáreas, los cuales han demostrado que la Amazonía occidental tiene comunidades muy ricas en especies de árboles, tal vez las más ricas del mundo (Gentry 1988, Valencia et al. 1994). También la cantidad de especies reportada en este estudio está en línea con las cifras elevadas reportados en dos florulas de la zona de Iquitos (Spichiger et al. 1996, Vázquez 1997), los cuales contienen unas 2740 especies.
En una escala regional, es más difícil comparar la diversidad de especies entre regiones, ya que no existe una forma de muestreo estandarizada, como en la escala local, donde las parcelas son de 1 - 0,1 ha. Uno tiene que utilizar florulas y listados de especies que tratan áreas de diferentes tamaños y están basadas en intensidades diferentes de estudio. Además tales registros son escasos. Sin embargo, se puede concluir que las selvas de la Amazonía occidental son muy ricas en especies también a una escala regional. Por ejemplo, la zona de Iquitos aparece mucho más rica en especies que las selvas mesoamericanas. Toda la flora de la Isla de Barro Colorado (16 km2) en Panamá tiene 1320 especies de plantas, y la flora de la Estación Biológica La Selva (15,1 km2) en Costa Rica incluye 1668 especies de plantas vasculares (Gentry 1990). De estas especies, probablemente no más de la mitad representan árboles de un tamaño similar al que se estudió en el presente trabajo.
Igualmente, los registros de la costa ecuatoriana y de la Amazonía central y sudoccidental citan números de especies mucho más bajos que los presentados en la zona de Iquitos, La flora de río Palenque (17 km2), en la costa del Ecuador, incluye 1033 especies; la Reserva Ducke (100 km. 2) cerca de Manaus en Brasil, cita 825 especies, y de Cocha Cashu en Madre de Dios se conocen 1856 especies. Estas cifras incluyen todas las plantas (Gentry & Dodson 1987, Gentry 1990), y por eso las cifras de especies para árboles son mucho menores que en la zona de Iquitos. En estas comparaciones es importante notar que las áreas mesoamericanas son muy bien estudiadas, y por eso no se espera un mayor aumento en el número de especies, mientras que las otras floras son casi seguramente muy incompletas (Gentry 1990).
Dentro de la Amazonía occidental es difícil a decir algo sobre los patrones geográficos en la riqueza de especies. En toda la Amazonía ecuatoriana (aproximadamente 71000 km2, hasta la altura de 600 m sobre el nivel del mar) se ha registrado la presencia de unas 3100 especies (Renner et al. 1990), y en toda la Amazonía peruana (aproximadamente 550 000 km2 hasta la altura de 500 m sobre el nivel del mar) se han observado ca. 7000 especies (Braco & Zarucchi 1993). Es muy probable que en ambas áreas se van a encontrar todavía muchas especies más.
Es notable que las cifras citadas arriba se refieren a todas las plantas y a superficies mucho más grandes que las 2,25 ha (0,0225 1=~) inventariadas en el presente estudio, donde se encontraron 1416 especies de árboles. En comparación con las otras cifras, esto realmente parece un número elevado. żSerá posible que un área tan pequeña de la Selva amazónica del Perú, donde sólo se contaron árboles, pueda contener casi un 50% del número de especies de plantas de toda la Amazonía ecuatoriana que representa una superficie de más de 3 millones de veces mayor? Probablemente la explicación a esta discrepancia está en la falta de colecciones y trabajo del campo en Ecuador. Además, hay que tomar en cuenta que las 3100 especies de Ecuador incluyen sólo plantas identificadas hasta la especie, no colecciones botánicas que no se han podido identificar. En cambio, nuestro estudio incluye casi 740 especies que son identificadas como morfoespecies pero no han podido referirse a un nombre específico. Este factor ha elevado el número de especies del presente estudio en relación a lo referido en la Amazonía ecuatoriana.
Otros estudios cuantitativos, en escalas comparables a nuestros inventarios en la zona de Iquitos, han producido números de especies comparables. Un inventarío en la zona del río Caquetá, Colombia, que incluyó 95 parcelas de 0,1 ha, donde se registraron todos los árboles mayores de 10 cm DAP, llegó a un total de 1077 especies con 7223 individuos (Duivenvoorden 1994, Duivenvoorden & Lips 1995). La riqueza de especies parece un poco menor que en la zona de Iquitos, pero eso puede ser un efecto del tamaño diferente utilizado en la definición de un árbol, ya que en otro estudio, Duivenvoorden (1994) ha demostrado que la mayor riqueza de especies se encuentra en los árboles menores de 10 cm DAP.
En la riqueza de especies, nuestros sectores de estudio presentaron valores más ó menos semejantes entre sí, con la excepción de los sectores en arena blanca los que tuvieron claramente menos especies que los otros sectores. Esta observación está en línea con los resultados anteriores (Anderson 1981, Clinebell et al. 1995, Duivenvoorden 1994) que también han demostrado que las zonas de arena blanca en la Amazonía contienen menos especies que los bosques en otros substratos.
Hay varias alternativas para explicar la baja diversidad en arena blanca. Se puede pensar que arena blanca es un substrato tan extremo que la misma existencia del bosque ya está en su límite, y por eso no hay tantas especies que hayan podido adaptarse a estas condiciones. También se ha sugerido (Connell & Lowman 1989) que en substratos como arena blanca donde los nutrientes son escasos, los árboles que forman en sus raíces un cierto tipo de simbiosis con hongos (ectomicorrhiza) ganan en competencia a otros árboles. Esto puede resultar en que los árboles sin ectomicorrhiza casi desaparecen de la comunidad arbórea, y consecuentemente el número total de especies disminuye. Además es posible que el número de especies especializadas a un cierto tipo de substrato depende de la superficie total disponible del substrato. Si es así, el bajo número de especies en arenas blancas puede simplemente ser el resultado de que las arenas blancas cubren áreas relativamente pequeñas, y no hay tanto espacio para la sobrevivencia de especies de árboles especializados como en otros tipos de suelo (MacArthur & Wilson 1967).
Lamentablemente es casi imposible tratar de evaluar cuál o cuáles de las alternativas explican mejor el tema. La primera alternativa, con seguridad explica por lo menos una parte de la incógnita. Es muy evidente que en la zona de Iquitos, en casos extremos de pobreza de nutrientes o de drenaje impedido, la arena blanca ya no puede soportar una vegetación de bosque, sino una vegetación baja de arbustos (Encarnación 1985, Ruokolainen & Tuomisto 1993). Sobre la segunda hipótesis de los ectomicorrhizas es muy difícil decir *algo, porque de ninguna especie de árbol, ni en arena blanca ni en otros substratos, se sabe que tipo de simbiosis posiblemente tienen con hongos. La tercera explicación puede ser aplicable, porque claramente las áreas de arena blanca son restringidas y no cubren superficies grandes. Por otra parte, hay que reconocer también que el conocimiento sobre la misma existencia, y aún menos sobre las extensiones de otros tipos de bosque que pueden ser ecológicamente relevantes, es todavía incipiente.
VARIACIÓN EN LA COMPOSICIÓN FLORÍSTICA Y SU CONEXIÓN CON LA VARIACIÓN AMBIENTAL
La variación en la composición de especies en los tres grupos de plantas estudiados en el presente estudio - árboles, Melastomatáceas y Pteridófitas - está correlacionada con la variación en las características de los suelos. Esto significa que, por ejemplo, si la flora indica que dos lugares son semejantes entre si pero un tercer lugar es diferente, entonces es bien probable que el suelo va a tener la misma tendencia de semejanzas y diferencias entre estos tres lugares. Hay que notar que la correlación no es al cien por ciento perfecta, lo que significa que a veces hay discrepancias en las formas como la flora y los suelos producen el patrón de semejanzas y diferencias entre los lugares. Sin embargo, es interesante que los tres grupos de plantas parecen estar más fuertemente correlacionadas con las mismas características del suelo: el contenido de arena y las concentraciones de calcio, magnesio y potasio.
La correlación entre la flora y los cationes es relativamente fácil de entender, porque estos son componentes importantes en el funcionamiento fisiológico de las plantas. Es interesante notar que el cuarto catión, sodio, no tuvo mucha participación con la variación florística, pero también su significado fisiológico para las plantas es menor que la de los otros cationes. Puede ser que la correlación observada entre la flora y el contenido de arena no es nada más que una manifestación de la misma influencia de los cationes, porque el contenido de arena está correlacionado en el área de estudio con la concentración de los cationes. Arena pura no tiene nutrientes disponibles para las plantas; los nutrientes en el suelo se encuentran mayormente en la fracción arcillosa. El contenido de arena es también una característica relacionada con el drenaje del suelo, porque la arena no tiene capacidad de retener mucha agua. Por lo tanto, después de las lluvias los suelos arenosos son más rápidamente drenados que los suelos arcillosos y consecuentemente son, generalmente, substratos más secos.
El fósforo, aunque es un nutriente importante para las plantas, no tuvo una conexión muy fuerte con la variación florística. En principio esta observación puede significar que en los sectores estudiados no hay mucha competencia por fósforo, que las plantas tienen satisfecha su demanda, o que no hay muchas diferencias entre las especies en las capacidades de extraer fósforo del suelo. La falta de competencia no parece muy probable, porque la concentración de fósforo es muy baja en todos los sectores. Tal vez, se puede pensar que esta concentración es tan mínima, que en términos prácticos tiene un significado casi nulo para cualquier especie. De esta manera podría ser que en efecto no hay muchas diferencias entre las especies. 0 tal vez que las plantas se abastecen de fósforo con métodos que no son directamente dependientes de las concentraciones en el suelo mineral: utilizan el fósforo que se libera en la mineralización de material orgánico, quizás con la ayuda de micorrhizas. En relación con el fósforo, y con las concentraciones de otros elementos también, es que los análisis químicos que se hace no necesariamente miden las concentraciones de una manera perfecta para entender cuanto de ese recurso se encuentra realmente disponible para las plantas en el suelo. En los análisis de laboratorio la extracción de los elementos de las muestras del suelo está basado en reacciones químicas que casi seguramente no son las mismas que ocurren cuando una planta está extractando elementos del suelo.
Las correlaciones entre pH y la flora, y entre LOI y la flora, resultaron relativamente débiles. Con los árboles existió una correlación estadísticamente significativa, pero con las Melastomatáceas y Pteridófitas no. Sin duda, el pH es una característica del suelo con mucha importancia para las plantas ya que afecta la disponibilidad de nutrientes, pero es posible que entre los sectores de estudio en el presente caso no había suficientemente variación para que las plantas hubiesen podido responder al pH en maneras detectables. El LOI se midió para obtener una idea de la cantidad de material orgánica, que es una característica importante en la estructura del suelo y en el bienestar de la microfauna y microflora (véase Capítulo 5), tanto como en su capacidad de retención de nutrientes y agua. Pero es posible que en el presente muestreo faltaba variación suficiente en LOI para producir correlaciones significativas con la flora. También hay que notar que LOI no es una medida exacta del contenido de material orgánico, porque durante la calefacción de la muestra no sólo sale carbón, sino también agua del cristal.
El aluminio extractable, en concentraciones grandes, es tóxico para muchas especies de plantas. En climas húmedos tropicales es común que los suelos contengan bastante aluminio, y por eso se podría pensar que durante su evolución la flora natural de este ambiente habría desarrollado resistencia a las concentraciones elevadas. El resultado de este estudio determina que hay una correlación, aunque no muy fuerte, entre aluminio y la variación florística, lo que sugiere que el efecto tóxico de aluminio es relevante para la flora natural. Por otra parte, la concentración de aluminio se correlaciona bien con el pH, porque el aluminio es uno de los factores que determinan el pH. Por eso es prácticamente imposible diferenciar los posibles efectos de estas dos características.
EFECTO DE LA DISTANCIA GEOGRÁFICA EN LAS SIMILITUDES FLORÍSTICAS
Es importante notar que la distancia geográfica entre los sectores de estudio no tuvo mucha importancia en la composición florística. Es relevante pensar en el posible efecto de la distancia geográfica, porque los lugares geográficamente más cercanos entre si deberían ser los más semejantes en su composición de especies, en el caso que no hubieran diferencias significativas en el suelo o en otras características físico-químicas en el medio ambiente. En tal situación, los patrones florísticos reflejarían mayormente la historia de la dispersión de las semillas o esporas y/o la historia de enfermedades, herbivoría, uso anterior por humanos, u otros disturbios bióticos o físicos en el bosque. Las bajas correlaciones entre las matrices florísticas y geográficas indican que la historia no ha tenido una importancia decisiva para determinar la composición de especies, o por lo menos es menor que la importancia de algunas de las características ambientales.
El énfasis en el papel de la historia llega a su extremo en la hipótesis que, en efecto, no es posible predecir la composición florística en un lugar en base a sus características ambientales (Hubbell & Foster 1986, Condit 1996). Según esta hipótesis, unas especies desaparecen y otras aparecen según las casualidades de la historia, sin control del medio ambiente.
La correlación relativamente alta que se observó en las distribuciones de especies entre los diferentes grupos de plantas en el presente estudio está demostrando que tiene que existir uno o más factores comunes que explican estos patrones semejantes. Es difícil creer que alguna enfermedad u otro factor biótico habría podido crear este patrón, ya que las plantas observadas pertenecen a grupos filogenéticamente lejanos. Tal vez, los patrones pueden explicarse con la recuperación de la flora después de un disturbio físico de gran escala, como un huracán muy grande, o un cambio fuerte en el clima (Prance 1982). Disturbios de este tipo pueden crear un patrón en la flora que se correlacione con la distancia geográfica. También algún tipo de barrera para la distribución libre, como una cadena de montañas o un cuerpo grande de agua, podría crear tales patrones. Pero la falta o bajo grado de correlación entre la flora y la distancia geográfica observada hace que este tipo de explicaciones sean muy improbables. Además, el río Amazonas con su planicie de inundación, es la única formación geográfica en la zona que podría actuar como barrera para la distribución de semillas, pero es muy fácil ver en las ordenaciones florísticas que las similitudes florísticas entre los sectores de estudio no dependen de su localización en el mismo lado del río o en el opuesto.
Con todo esto no queremos decir que el conocimiento de la historia no sea potencialmente importante para entender la variación en la composición de especies entre diferentes lugares. Por ejemplo, en nuestro estudio es probable que los sectores Momón 1 y Momón 2 hayan sido afectados más que los otros sectores por tumba selectiva de madera, cosecha de productos no-maderables y otras actividades humanas. Además, hay que hacer notar que la falta de correlación entre las matrices de similitud florística y de distancia geográfica sólo excluye efectos que presentan una gradiente geográficamente continua. Factores históricos que aparecen esporádicamente en manchas separadas, por ejemplo la sucesión del bosque iniciado en claros creados por caídas de árboles o después de abandono de chacras aisladas, no pueden ser detectados con este análisis. Aunque en la selección de las parcelas para el muestreo de los árboles se trataba de evitar claros recientes para disminuir los efectos de sucesión en los resultados, es posible que las parcelas mostraban en realidad diferentes fases de sucesión más tardía.
EFECTO DE GEOLOGÍA EN LA COMPOSICIÓN FLORÍSTICA
Si se piensa en la historia a una escala mayor, como en términos de procesos geológicos, se ve que en efecto la historia puede explicar bastante sobre las diferencias y semejanzas físicas y químicas en los suelos (véase Capítulos 4 y 12 para una discusión más detallada). Según su historia geológica, los sedimentos superficiales pueden haber tenido originalmente composiciones físicas y químicas muy variables. Al pasar el tiempo, los procesos de formación de suelos (véase Capítulo 5) pueden modificar bastante a las características de los sedimentos, pero muy a menudo sobreviven rastros de su origen, los cuales son importantes para la vegetación.
Por ejemplo, nuestros 32 sectores de estudio pueden clasificarse en seis grupos según la composición de especies de pteridófitas o Melastomatáceas, y esta clasificación es relativamente fácil de explicar en términos geológicos. Los sectores en el grupo 1 (véase Figura 7.8), con las más altas concentraciones de cationes y con textura arcillosa de los suelos, representan áreas donde los sedimentos más superficiales han sido depositados en un ambiente semimarino durante el Mioceno tardío (Formación de Pebas, Capítulos 4 y 12). Los sectores en el grupo 2 (Carbajal km0 y km2, San Antonio 1) están caracterizados por sedimentos depositados en un ambiente parecido al anterior, pero con menos influencia marina (Unidad B). Los sectores del grupo 4 (San Antonio 2, Nauta, Carbajal km4, km8, kml0 y kml2) se encuentran en áreas cubiertas por sedimentos aún más jóvenes, pero todavía con algunos rastros de mareas (Unidad C). El grupo 5 (Maniti, Mishana 1, Momón 1, Panguana 1 y 2, Libertad Agraria km3, San Pedro) incluye todos los sectores situados en terrazas relativamente recientes de diferentes ríos. El grupo 6 incluye todos los sectores con sedimentos arenosos que han sido reciclados varias veces por los ríos.
El único grupo de sectores que no tiene una correspondencia simple a la geología es el grupo 3. De los sectores pertenecientes a este grupo, Momón 2 tiene sedimentos que mineralógicamente parecen tener clara influencia marina, pero la concentración de cationes es más baja de lo que se esperaría en este tipo de sedimentos (Tabla 12.2 en el Capítulo 12). Una posible explicación para esta discrepancia, es que este sector puede haber sido cultivado algún tiempo atrás, lo cual puede haber agotado las reservas de nutrientes en los suelos. También Santa Ana y 13 de Febrero arcilla presentan características mineralógicas que indican influencia marina, pero tal vez un poco menos que en el caso de Momón 2. Además, 13 de Febrero arcilla queda contiguo a un área con suelos arenosos, y por eso es posible que allí se encuentren especies que en realidad no prefieren ese tipo de suelo, pero crecen allí simplemente porque hay flujo abundante de sus propágulos desde las áreas de arena. Tal efecto de la dispersión hace naturalmente más difícil la detección de una posible relación entre flora, suelo y geología. El último sector de este grupo, Palo Seco 2, no ha sido analizado mineralógicamente. Tiene suelos relativamente arenosos y pobres en nutrientes, y en verdad representa una mancha arenosa rodeada por áreas con suelos arcillosos que mayormente están bajo cultivo. Por eso, puede ser que también en este sector haya influencia de dispersión de algunas especies desde un área edáficamente diferente, lo cual resulta en una situación difícil de explicar en la ordenación.
COMPARACIÓN DE LOS PATRONES FLORÍSTICOS ENTRE LOS DIFERENTES GRUPOS DE PLANTAS
Tanto las ordenaciones como los análisis de correlación indican claramente que árboles, Melastomatáceas y Pteridófitas dan una imagen bien semejante de las relaciones florísticas entre diferentes sectores de estudio. Especialmente la correlación entre árboles y Pteridófitas es muy fuerte. Los patrones florísticos según Melastomatáceas defieren algo más de los patrones basados en los otros dos grupos de plantas. En comparación con los árboles, los datos de las Melastomatáceas colocan a los sectores Momón 1 y Momón 2 de manera un poco distinta, lo cual puede ser relacionado con la pequeña superficie muestreada, y con los disturbios humanos en estos sectores. Entre pteridófitas y Melastomatáceas, son solamente los sectores con suelos pobres y arenosos que salen diferentes en las ordenaciones. Esto casi puede explicarse, por la cantidad menor de Melastomatáceas especializadas en crecer en arena blanca, prácticamente sólo Clidemia epibaterium. Consecuentemente, en tales lugares la mayoría de las pocas especies de Melastomatáceas que se encuentran son generalistas, pudiendo crecer en lugares ecológicamente muy diferentes. Por lo tanto, las Melastomatáceas no sirven tan bien como las Pteridófitas para diferenciar entre tipos florísticos del bosque en los suelos más pobres.
A pesar de algunas diferencias, las semejanzas en los patrones florísticos entre los tres grupos de plantas son tan fuertes que es bien posible utilizar cualquiera de los grupos como un indicador del comportamiento de los otros. Estos tres grupos de plantas no incluyen toda la flora, pero en todo caso hasta más de la mitad de las especies (Gentry 1990, Duivenvoorden 1994, Vázquez 1997, Balslev et al. en prensa), y por eso es justificable sugerir que las Melastomatáceas y/o Pteridófitas pueden ser utilizadas para indicar patrones generales para toda la flora. Las posibilidades y restricciones en el uso de estas plantas como indicadoras de diferentes tipos de bosque está tratado con más detalle en el Capítulo 13.
RELACIÓN ENTRE IMÁGENES DE SATÉLITE Y COMPOSICIÓN FLORÍSTICA
La buena correlación entre los diferentes grupos de plantas es sin duda la mayor razón para explicar la correlación que se observa entre las Melastomatáceas y Pteridófitas por un lado, y los patrones de reflectancia de luz medidos por el satélite Landsat TM por el otro. Obviamente el satélite no puede detectar directamente estas plantas, ya que crecen mayormente en el sotobosque y no llegan hasta el dosel. La variación en los valores de reflectancia es más que todo causada por diferencias en la estructura y coloración de copas, ramas y hojas de los árboles, y tal vez por la actividad fotosintética. Estas características estructurales y funcionales corresponden a diferencias en la composición de especies de árboles, lo cual por su parte tiene una correlación con las diferencias en la composición de especies de Melastomatáceas y Pteridófitas. De esta manera, la conexión entre las imágenes de satélite y los patrones florísticos en Melastomatáceas y Pteridófitas es indirecta. Por eso, se puede considerar esta conexión como una indicación de un nivel relativamente alto de organización predecible en la vegetación.
Aunque existe una correlación entre la variación florística y los patrones de reflectancia de luz de la imagen de satélite, parece que en esta correlación hay diferencias regionales dentro del área de estudio. Una discrepancia notable entre los patrones florísticos y los de la imagen de satélite es que la imagen de satélite hace una diferencia entre los dos lados del río Amazonas, pero la flora no lo hace. Sin embargo, dentro de cada una de las regiones separadas por el río, la correlación entre la flora y los patrones de la imagen de satélite parece relativamente buena. Este resultado sugiere que las imágenes de satélite pueden utilizarse efectivamente para reconocer y cartografiar aproximadamente los diferentes tipos de bosque, pero que en las imágenes de satélite posiblemente existen algunos patrones regionales o de gran escala que no son directamente transferibles a la flora. Una explicación tentativa es que el lado oriental del río Amazonas realmente tiene suelos generalmente más pobres que el lado occidental, y que los sectores 7 de Julio y Libertad Agraria km8 representan manchas muy pequeñas de suelos bien arcillosos y con concentraciones relativamente altas de cationes. Para los análisis de correlación, las reflectancias de luz fueron medidas en una superficie más grande que la del muestreo botánico, y por lo tanto es posible que cuando se calcularon los promedios de valores de reflectancia, se incluyeron varios Pixeles que en la realidad representan áreas florística y edáficamente diferentes. Es obvio que se necesitan todavía más estudios para aclarar la conexión entre los tipos de bosque y las imágenes de satélite (Tuomisto et al. 1995).
CLASIFICACIÓN DE LOS BOSQUES
Aquí cabe mencionar que el concepto 'tipo de bosque' es una abstracción humana con el fin de describir y entender la variación en la composición florística y de sus determinantes ambientales, aunque estas variaciones en realidad pueden ser continuas. A veces es posible observar áreas grandes aparentemente homogéneas, que son limitadas por zonas donde las características florísticas y ambientales cambian rápidamente en distancias cortas. En tales casos tiene mucho sentido hablar de tipos diferentes de bosque. Pero muchas veces los cambios pasan gradualmente y sin límites obvios, de manera tal que los tipos definidos no son más que unas secciones de una gradiente continua, y se encuentran fácilmente lugares que son intermedios entre los tipos definidos y por lo tanto difíciles de clasificar. Una de las tareas para los futuros estudios es justamente investigar hasta que punto las variaciones en la flora y en el medio ambiente son graduales, y hasta que punto es útil tratar de clasificar la variación entre los extremos.
El número de tipos de bosque que uno puede diferenciar a lo largo de la gradiente florística depende mucho de la decisión subjetiva del investigador, y del número de lugares inventariados. Por ejemplo, los 15 sectores del presente estudio donde se hicieron inventarío de árboles, justifican el agrupamiento en cuatro grupos, o sea, el reconocimiento de cuatro tipos de bosque. Pero cuando se consideran los 32 sectores donde había muestreo de Melastomatáceas y Pteridófitas, parecería más apto identificar seis tipos diferentes, aunque los extremos de la gradiente observada fueran los mismos. Generalmente, cuando el estudio incluye más lugares, pueden distinguirse diferencias más finas a lo largo de la gradiente. Pero el aumento de observaciones también hace posible estimar si en la variación de la flora y del medio ambiente hay discontinuidades que pueden justificar la definición de tipos de bosque.
Las investigaciones realizadas indican que la flora en los bosques de tierra firme en la zona de Iquitos está compuesta hasta cierto punto de una manera predecible, ya que las distribuciones de la mayoría de las especies de plantas reflejan los mismos factores ambientales e históricos. Entre los factores edáficos más importantes están la textura de los suelos y sus contenidos de calcio, magnesio y potasio; los patrones florísticos tanto de los árboles como de las Melastomatáceas y Pteridófitas están correlacionados con estos factores. La variación de los suelos está relacionada con la historia geológica, de la cual depende los tipos de sedimentos que existen en cada lugar, y la magnitud del cambio ocasionado por procesos de temperización y formación de suelos.
Las notables diferencias florísticas, identificadas entre los bosques en diferentes sitios, sugieren que para el manejo y la utilización sostenibles de los recursos biológicos es importante y urgente el reconocimiento y el mapeo de tales diferencias. Sólo de esta manera se puede garantizar que cada sitio sea manejado de acuerdo a sus características naturales, ya sea para fines agropecuarios, forestales o de conservación. La altísima riqueza de especies de plantas en los bosques de tierra firme de la zona de Iquitos es un recurso valioso, pero también difícil de cuantificar por las dificultades en identificar las especies. Por esta razón, no es factible basar el mapeo de los tipos diferentes de bosque en toda la flora. Los resultados del presente estudio sugieren que las Melastomatáceas y pteridófitas pueden servir como indicadoras de patrones florísticos más generales en la zona, y tal vez también en otras partes de los bosques húmedos de la Amazonía.
También los patrones visibles de imágenes de satélite Landsat TM pueden indicar diferencias y semejanzas entre la vegetación de diferentes lugares, y pueden ser utilizadas para la identificación y el mapeo de los tipos diferentes de bosque. Esto es sugerido por la correlación observada entre la variación geográfica en las reflectancias de luz en la imagen de satélite por un lado, y en las composiciones florísticas de los árboles, las Melastomatáceas y las Pteridófitas por el otro lado. Cabe mencionar, sin embargo, que falta todavía precisar más esta posibilidad, y en todo caso los estudios basados en imágenes de satélite siempre deben ser acompañados por verificación en el campo para evitar interpretaciones erróneas.
Este estudio demuestra que es posible hacer mapeos para los fines de mejor manejo y conservación de bosques amazónicos utilizando imágenes de satélite junto con inventarios de campo basados en el uso de helechos y Melastomatáceas como indicadoras de variación ambiental.
AGRADECIMIENTOS
Agradecemos la colaboración muy valiosa de todas las personas que nos acompañaron y apoyaron en el trabajo de campo, especialmente a Abel Sarmiento, Richer Ríos, Alberto Torres, Mildred García, Antonio Layche, Juan Ruíz, Melchor Aguilar, Simón Cortegano, Guillermo Criollo, Nestor Jaramillo, Gustavo Torres, César Bardales, Lizardo Fachín, Illich Arista, Isabel Oré, Jaana Vormisto, Elina Lusa, Hannele Koivunen, Juha Järvinen, Vesa Koivunen y Tomas Roslin. También agradecemos a las comunidades y personas que nos facilitaron el trabajo en Mishana, Tarapoto, Gengen, Libertad Agraria, San Antonio, Carbajal, 13 de Febrero, Santa Ana, Nauta (km9.5 km. de la carretera), Panguana 2a zona y San Pedro. Las Facultades de Ingeniería Forestal y de Biología de la Universidad Nacional de la Amazonía Peruana (UNAP) nos apoyaron de múltiples formas en el trabajo de campo y de gabinete. El Museo de Suelos de la UNAP nos facilitó una oficina. El Herbario Amazónico de la UNAP, el herbario de la Universidad de Turku y Rodolfo Vázquez nos prestaron apoyo logístico en el procesamiento de las muestras botánicas. José Torres nos ayudó de múltiples maneras. Muchos especialistas contribuyeron a las identificaciones de las muestras botánicas, especialmente Henrik Balslev, Cornelis C. Berg, Douglas C. Daly, Alwyn H. Gentry, Karl U. Kramer, Ron Liesner, Paul Maas, John T. MickeI, John Mitchell, Robbin C. Moran, Scott A. Mori, Terry Pennington, Susanne S. Renner, Alan R. Smith, Charlotte Taylor, Rodolfo Vázquez, Henk van der Werff y John Wurdack. El Instituto Nacional de Recursos Naturales nos proporcionó los permisos necesarios para colectar y exportar muestras botánicas.
