

UN ESTUDIO Y SACA EXPERIMENTAL DE Cebuella pygmaea*
![]()
(* Versión original presentada a Min. Agric. -Perú, en Mayo 1979).
Pekka Soini y María de Soini
INTRODUCCION
Con la finalidad de ampliar los estudios de ecología, dinámica poblacional y manejo de Cebuella pygmaea, llevados a cabo principalmente en la cuenca del Río Manití entre 1976 a 1978 (Soini, en prensa). Se realizó un estudio complementario en la misma área desde Diciembre 1978 a Febrero 1979. El presente estudio incluyó un segundo censo de la población de Cebuella y una saca experimental, habiendo capturado y examinado 115 ejemplares, que permitió evaluar los efectos inmediatos de la saca y analizar la estructura en detalle de lo que fue posible en el estudio anterior, aportando información indispensable para el manejo de este recurso natural. Los objetivos principales del estudio fueron: 1) realizar un segundo censo completo de la población; 2) analizar en detalle la estructura poblacional, 3) continuar las investigaciones sobre las características reproductivas y 4) estudiar las consecuencias demográficas de la saca en la población afectada.
Una descripción detallada del área y una caracterización de la organización de Cebuella fueron dadas previamente por Soini (1982).
MÉTODOS
Utilizando los procedimientos descritos anteriormente, se localizaron e identificaron todas las unidades sociales de Cebuella existentes dentro del área de estudio (Soini 1982).
Los métodos de captura y examen de los animales fueron los mismos empleados en Saguinus fuscicollis, y están descritos en el trabajo correspondiente (Soini, este volumen). Antes de proceder a la captura de una manada, ésta fue sometida a observación detenida, a fin de constatar el número total de animales y la condición reproductiva de la misma. Para registrar los efectos de la extracción de individuos en el comportamiento de la manada remanente, los integrantes no capturados fueron observados continuamente hasta completar la captura de la manada; cuando la saca fue sólo parcial, la observación post-captura de la manada residual fue continuada por una semana o más.
Clasificación de los animales por edad
La edad aproximada de los individuos fue estimada en base a los conocimientos previamente adquiridos sobre desarrollo físico y etológico de Cebuella (Soini, en prensa), complementado con la información existente sobre el desarrollo de Callithrix jacchus (Johnston et al. 1970, Abbot y Hearn 1978), especie filogenéticamente muy cercana a Cebuella (Hershkovitz 1977). Los individuos fueron clasificados en cuatro categorías principales de edad, como sigue:
Infantes: Individuos cuya edad fue estimada en menor de 0,5 año. Se caracterizan por su tamaño pequeño, cabeza desproporcionadamente grande y dientes M2 todavía no erupcionados. Dentro de esta categoría, las crías dependientes constituyeron una subcategoría que comprendió los infantes de dos meses de edad y que son frecuentemente cargados por los otros miembros de la manada.
juveniles: Individuos cuya edad fue estimada entre 0,5 - 1,0 año. Ligeramente más grandes que los infantes, pero de aspecto más delgado, dientes M2 presentes, pero los caninos permanentes aún sin erupción; pelaje negro en el circum ano-genital todavía incipientemente desarrollado.
Subadultos: Individuos cuya edad fue estimada entre 0,5 a 1,5 años. De menor tamaño que los adultos y generalmente de aspecto más delgado; pigmentación negra del escroto incipiente y vulva de las hembras despigmentada; caninos permanentes presentes.
Adultos: Individuos físicamente maduros, cuya edad fue estimada en 1,5 años o más. Los adultos jóvenes, entre 1,5 - 2 años de edad, fueron diferenciados de los adultos mayores por la condición impecable y sin desgaste de sus caninos y otros dientes, así como por su aspecto general más 'Juvenil".
RESULTADOS Y DISCUSIÓN
Tamaño y peso
El tamaño y peso de los animales examinados están en el Cuadro 1. En los adultos, las hembras son significativamente más grandes que los machos (Cuadro 2).
Cuadro 1. - Pesos y medidas de las diferentes categorías de edad de Cebuella pygmaea.
| Edad & Sexo | (N) | P.ESO (g) X ± DE |
Long.cuerpo (mm) X ± DE |
Long.oola(mm) X ± DE |
|
| Adulto | M | -36 | 110,1 ±11,0 | 135,2±4,7 | 197,8±8,6 |
| Adulta | H | -35 | 121,8 ± 9,9* | 140,8± 5,3 | 204,1 ±9,3 |
| Subadulto | m | -7 | 99,7±10,7 | 130,6±3,8 | 202,1±5,9 |
| Subadulta | H | -7 | 113,6±15,2 | 137,9±4,2 | 201,6±11,6 |
| juvenil | MyH | -22 | 63-90 | 111-130 | 173-200 |
| Infante2. | MyH | -14 | 42-66 | 97-119 | 142-188 |
| Infantel." | MyH | -10 | 27-47 | 76- 95 | 114-153 |
* Esta cifra incluye sólo a 27 hembras no preñadas de la muestra. La linclusión de las 8 hembras preñadas daría un peso promedio de 128,7 ± 16,7 g. La hembra preñada más pesada de la muestra pesaba 165 g.
** La muestra noincluye neonatos; el infante más joven
y pequeflo tenía 24 días de edad.
Cuadro 2. - Comparación de los pesos y algunas medidas corporales de los machos y hembras adultos de C. pygmea.
|
Medidas |
Sexo |
X D.E. |
N |
t P" |
| Peso (g) | M | 110,1± 11,0 | 36 | 4,362 < 0,0005 |
| H | 121,8*± 9,9 | 27 | ||
| Long. cuerpo (mm) | M | 135,2 ± 4,7 | 36 | 4,603 < 0,0005 |
| 0140,8 ± 5,3 | 30 | |||
|
Long. cola (mm) |
M |
197,8 ± 8,6 |
36 |
2,855 < 0,0005 |
|
204,1 ± 9,3 |
30 |
|||
| Long. rodilla | M | 46,7 ± 1,6 | 36 | 4,393 < 0,0005 |
|
Talón (mm) |
H | 48,4 ± 1,5 | 35 |
M = machos; H = hembras
* Las hembras preñadas fueron
excluidas del cómputo.
** Prueba Y para una cola.
Densidad poblacional
El censo completo dentro del área de estudio de 3 km2 reportó un total de 118 individuos de locomoción independiente. La densidad ecológica o densidad en el bosque de inundación fue de 59 ind./km2, con una biomasa concomitante de 6,2 kg/km2. El bosque de ribera constituye el óptimo hábitat de Cebuella (Soini 1982), cuya densidad fue de 227 ind./km2, con una biomasa de 26,1 kg/km2.
Comparando resultados del presente censo con los del censo efectuado hace un año en la misma área, indica un aparente incremento de 15% en la población total (Cuadro 3); mientras que la composición por categorías de edad, se mantuvo inalterada (Fig. 2). La diferencia encontrada entre los dos censos probablemente refleja una fluctuación normal y fortuita de manadas dentro de un área dada, aunque cabe la posibilidad que alguna manada fuera inadvertidamente omitida en el primer censo.
Proporción macho / hembra.
En este censo fue posible identificar el sexo del 93% de los individuos de locomoción independiente (Cuadro 4). Omitiendo los 8 individuos cuyo sexo no fue determinado, la proporción macho/hembra fue 55:55. Los resultados indican que la proporción macho/hembra se mantiene aproximadamente 1:1 en todas las categorías de edad.
Composición de Las manadas
Como se constató anteriormente (Soini 1982), las manadas reproductivas de Cebuella están formadas básicamente por una pareja reproductora y su prole de hasta cuatro camadas sucesivas (Algunas recientes observaciones de campo sugieren la posibilidad que el enlace entre la hembra reproductora y su macho no sería necesariamente permanente, sino que podrían suceder sustituciones de los machos dominantes. Naturalmente este hecho no cambiaría que, en cualquier momento dado, la manada estuviera integrada de una pareja reproductora, aunque todos los descendientes no serían necesariamente engendrados por el mismo). El análisis de la composición (Cuadro 4) demostró que la mitad de ellas contenía, además de la pareja reproductora, de uno a tres individuos adultos, siendo el promedio general de 2,8 (1,4 machos y 1,4 hembras) por manada. Del total, 72% eran adultos jóvenes, recientemente llegados a la madurez sexual, muchos de ellos aparentemente todavía no se habían desligado de sus manadas natales. Sin embargo, en otros casos, era evidente que se trataba de individuos asociados, que no formaban parte de la prole de la pareja reproductora, lo que indica que la presencia permanente o transitoria de adultos asociados en las manadas es un aspecto normal de la organización social en esta especie.
Estructura poblacional
La muestra poblacional,de 115 ejemplares, capturados dentro y alrededor del área de censo, abarca el 83% de la población total observada y ploteada en el mapa (Fig.1). Estadísticamente, la composición por edad y sexo de esta muestra no difiere significativamente de la composición de la población censada (test de chi-cuadrado para la bondad de ajuste : X2 = 2,82, con 4 grados de libertad, p < 0,50). Esto nos permite hacer inferencia más detalladamente sobre la estructura poblacional en base a la presente muestra.
Considerando que en los mamíferos silvestres el desgaste dentario lleva una buena correlación con la edad cronológica del animal (Taber 1971) clasificarnos a los adultos en cuatro niveles de edad (según el grado de desgaste de sus caninos) como sigue:
A = caninos intactos B = caninos un poco desgastados
Cuadro 3.- Resultados del censo poblacional de C. pygmaea en el área de estudio, Río Manití (Diciembre 1978 - Enero 1979) y comparación con los resultados del censo anterior.
| Censo de 1978 | Censo de 1977 | |
| Población total-* | 118 | 103 |
| Total manadas: | 21 | 16 |
| Total parejas incipientes: | 4 | 7 |
| Total solitarios: | 4 | 4 |
| Densidad ecológica: | 59 ind./krn2 | 51,5 ind./kn,2 |
| Densidad en la ribera:-* | 227ind./km2 | 210***ind./km2 |
| Biomasa ecológica: | 6,2 kg/kM2 | 5,3 kg/km2 |
| Biomasa en la ribera: | 26,1 kg/kM2 | 23,7kg/kM2_ |
* Las crías cargadas (= menores de 2 meses de edad) no están incluidas en los cómputos.
** Incluye todas las manadas e individuos que habitaban en el bosque de las orillas del Río Manití dentro del área de estudio. La densidad fue calculada a base de una faja continua de ocupación de 50 m de ancho, desde ambas orillas hacia el interior del bosque.
*** En Soini (1982) se aplicó un método de cómputo más preciso, basado en conocimientos más detallados sobre la extensión de las áreas ocupadas por las unidades sociales de C. pygmaea en el bosque de ribera, dando por densidad 247 ind./km2; Sin embargo, aquella cifra no es directamente comparable con el resultado segundo censo.
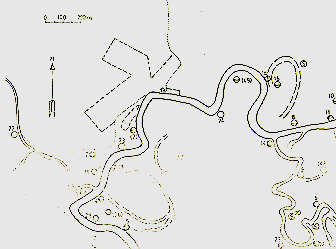
Fig. 1.- Mapa del área de estudio
en "Los Angeles", Río Manití. El rectángulo delimita
el área de censo de C. pygmaea. Los círculos numerados
indican las ubicaciones de las manadas y otras unidades sociales de C. pygmaea
presentes en el área de estudio en Diciembre 78 - Enero 79 (círculos
con el número de referencia en paréntesis indican parejas incipientes
y círculos con letra S inserta, solitarios). Las unidades sociales que
fueron sometidas a saca total están señaladas con círculo
sombreado, las de saca parcial con círculo semi-sombreado y las dejadas
intactas con círculo blanco.
C = caninos regularmente desgastados
D = caninos muy desgastados 1
La Fig. 3 presenta el perfil poblacional construido en base a la muestra representativa de la población. La forma del perfil nos sugiere que la mortalidad acumulativa afecta a ambos sexos en igual proporción.
Reproducción
Aun cuando es,posible observar ocasionales camadas en cualquier mes del año, Cebuella presenta dos temporadas principales de nacimientos, que oscilan alrededor de los meses Diciembre y Junio, (Soini 1982). Observaciones durante el presente estudio señalan que 87% de las manadas estaban reproductivamente activas en el período Octubre a Febrero* (Una manada fue considerada reproductivamente activa dentro de la temporada, si la hembra dominante estaba lactando crías nacidas después de Setiembre o si la hembra estaba preñada y próxima a parir antes de fines de Marzo) (Cuadro 5). Esta tasa de reproducción es idéntica a la observada en la misma localidad y temporada de hace dos años atrás (Soini 1982), lo que indica que la tasa de nacimientos se mantiene constante en la población.
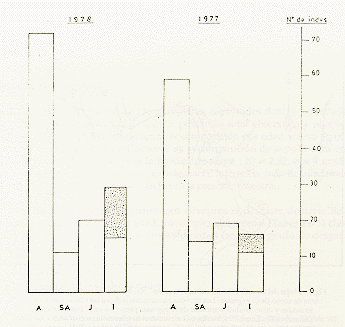
Fig. 2.- Composición de la población
de C. pygmaea en el presente censo
y en el censo realizado un año antes. La porción punteada refiere
a crías cargadas.
El 80% de las camadas de crías dependientes
o de fetos abortados consistían de dos crías o fetos/parto (Cuadro
5), que confirma las observaciones anteriores, donde casi todas las camadas
en estado natural consistieron en mellizos.
Cuadro 4.- Composición de la población de C. pygmaea
en el área de estudio, Río Manití (Diciembre 1978 - Enero
1979).
|
Unidad Social |
Adultos |
adultos |
Subadultos |
Juveníles |
Infantes |
Crías |
Total* |
||||||||||
|
M |
H |
M |
? | H |
M |
? | H |
M |
? | H |
M |
H |
M |
? |
H |
||
|
1a |
1 |
2 |
2 |
5 |
|||||||||||||
|
1b |
1 |
1 | 2 |
2 |
|||||||||||||
|
3 |
1 |
1 | 1 | 3 | |||||||||||||
|
4 |
1 |
1 |
1 |
1 | 2 | 1 | 7 | ||||||||||
| 5 | 1 | 1 |
|
2 |
1 |
1 | 1 |
|
2 | 7 | |||||||
| 6 | 1 | 1 | 1 | 1 | 4 | ||||||||||||
|
7 |
1 |
1 | 1 | 1 | 4 | ||||||||||||
|
8 |
1 |
1 | 1 | 1´ | 1 | 1 | 8 | ||||||||||
|
9 |
1 |
1 | 1 | 1 | 2 | 4 | |||||||||||
|
10 |
2 |
1 |
|
2 | 2 |
2 |
7 | ||||||||||
|
11 |
1 |
1 |
1 |
1 | 3 | ||||||||||||
|
13 |
1 |
2 | 1 | 1 | 1 | 1 | 1 | 8 | |||||||||
|
14 |
1 |
1 | 1 | 3 | |||||||||||||
|
17 |
1 |
1 |
|
1 |
1 |
2 | 1 | 1 | 6 | ||||||||
|
19 |
1 |
1 | 1 | 3 | |||||||||||||
|
22 |
2 |
1 | 2 | 5 | |||||||||||||
|
23 |
2 |
1 | 1 | 1 | 1 | 6 | |||||||||||
|
24 |
1 |
1 | 1 | 1 | 1 | 5 | |||||||||||
|
25 |
1 |
1 | 1 | 3 | |||||||||||||
|
27 |
1 |
1 | 1 | 1 |
|
2 | 5 | ||||||||||
|
28 |
1 |
1 |
1 |
1 | 1 | 1 | 1 | 7 | |||||||||
| Parejas incipientes | |||||||||||||||||
|
2 |
1 |
1 |
|
2 | |||||||||||||
|
26 |
1 | 1 |
1 |
2 | |||||||||||||
|
48 |
1´ |
1´ |
|
1 | 2 | ||||||||||||
|
49 |
1 |
|
1 | 3 | |||||||||||||
| Solitarios | |||||||||||||||||
|
19a |
|
1 | 1 | ||||||||||||||
|
20 |
1 |
1 | |||||||||||||||
| 13 | 1´ |
|
|
1 | |||||||||||||
| 22 | 1´ | 1 | |||||||||||||||
| Total | 28 | 25 | 7 | (3) | 9 | 5 | (3) | 3 | 8 | (2) | 10 | 7 | 8 | 3 | (9) | 2 | 118 |
* No incluye las crías cargadas
M= macho;? =sexo desconocido;
H =hembra; V=determinación dela edad tentativa
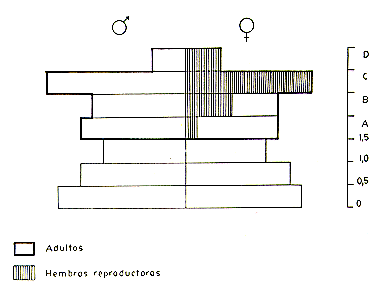
Fig. 3.-Perfil poblacional de C. pygmaea
(para la definición de los niveles de edad ver el texto).
De las hembras sexualmente maduras de la población, 63% estaban reproductivamente activas (preñadas, lactantes o post-lactantes). Su distribución por categorías de edad dentaria (Fig. 3) indica que todas las hembras adultas no-reproductivas pertenecen a las dos categorías de menor edad y que todas las hembras reproductoras pertenecen a las dos categorías de mayor edad. La ausencia de hembras de mayor edad que no hayan reproducido parece indicar que todas las hembras sobrevivientes y normales alcanzan eventualmente la posición de reproductora (hembra alfa) en la población.
Proporción adulto/inmaduro y sus implicaciones en el manejo
Los adultos comprendieron el 54,5% de la población censada. Un año antes, la proporción fue de 54,6% (Soini 1982), lo que indica que la proporción adulto/inmaduro se mantenía constante. Un mayor desvío probablemente significaría que la población no se encuentra demográficamente estabilizada, que podría ser debido a disturbios causados por factores climatológicos y/o intervenciones del hombre. Si en una población estabilizada la proporción se mantiene aproximadamente constante, donde los nacimientos y la mortalidad se mantienen en equilibrio, podremos valemos de este parámetro para evaluar la condición demográfica de una población. De acuerdo a la observación hecha en la población estudiada, que aparentemente se encuentra estabilizada, la proporción adulto/inmaduro en una población estabilizada debería ser aproximadamente 1:0,85. Para determinar la proporción adulto/inmaduro en una población, sería necesario ubicar los individuos en el área, incluyendo los solitarios y las parejas incipientes, que son más difíciles de ubicar. En vista de la magnitud de esfuerzo y tiempo requerido para un censo completo de una población, sería más práctico ignorar los solitarios y parejas incipientes, y utilizar como parámetro únicamente la proporción adulto/inmaduro. Para averiguar esta proporción sería necesario ubicar y censar algunas manadas de muestreo. En la Población de estudio, la proporción adulto/inmaduro es en término medio 1:1,1.
Cuadro 5. - Estado reproductivo de
las manadas de C. pygmaea en el área de estudio.
| Manada N° |
Estado Reproductivo* |
Fecha Parición** |
Tamaño Camada*** |
Sexo**** | |
| 1a | C | Octubre | 2 | M | M |
| 1b | C | Noviembre | 2 | ||
| 3 | P | Febrero | (?) | ||
| 4 | P | Enero | (2) | M | M |
| 5 | C | Octubre | 2 | M | M |
| 6 | - | ||||
| 7 | P | 12.01 | 2 | ||
| 8 | ? | ||||
| 9 | C | Diciembre | 2 | ||
| 10 | C | Enero | 2 | ||
| 11 | C | Diciembre | 2 | M | H |
| 12 | C | Noviembre | 1 | H | |
| 13 | P | Febrero | (2) | ||
| 14 | P | 19.01 | 2 | M | H |
| 16 | C | Octubre | M | ||
| 17 | C | Octubre | 2 | H | |
| 18 | C | Enero | 1-2 | ||
| 19 | P | 3.02 | 1-2 | H | |
| 21 | ? | ||||
| 22 | - | ||||
| 23 | P | Febrero | (2) | M | H |
| 24 | ? | ||||
| 25 | - | ||||
| 27 | C | Diciembre | 2 | ||
| 28 | P | ? | |||
| 29 | C | Diciembre | 2 | ||
| 30 | ? | ||||
* Estado de la hembra reproductora cuando fue capturada u observada (C = con crías lactantes; P - preñada; ? = estado reproductivo no determinado; - = sin crías lactantes ni preñada).
** Fecha o mes de parición observada o estimada.
*** Número de crías observadas o fetos [ = 01 abortados.
**** Sexo de las crías o fetos examinados (M = macho; 11 = hembra).
Saca experimental
Desde Diciembre de 1978 a Febrero de 1979, capturamos y examinamos
115 individuos, pertenecientes a 28 unidades sociales. De éstos, cinco
fueron soltados, dos fugaron luego del examen y dos murieron en la estación
de retención del Proyecto en Iquitos. 83% de los individuos fueron capturados
dentro del área de censo (3 Km2) y los 29 restantes provinieron de cuatro
manadas situadas en la periferia del área de estudio (Cuadro 6, Fig.
D. A fin de observar las consecuencias demográficas de la saca, el área
de estudio fue dividida en tres zonas sujetas a diferentes intensidades de saca,
separadas entre sí por cursos de agua
(Fig. 4).
Cuadro 6.- Resultados dela saca de C. pygmaea
en el área de estudio. Las unidades sociales que estaban ubicadas
fuera del área de censos están denotadas en paréntesis.
|
N° de ref |
Fecha |
Cosechados |
N° cosechados |
Total |
|||||||||
|
Adultos |
Inmaduros |
Adultos |
Inmaduros |
||||||||||
|
M |
H |
M |
H |
M |
? |
H |
M |
? |
H |
||||
| 1a | A | 22-12 | 2 | 1 | 2 | 5 | |||||||
| 1b | A | 22.12 | 1 | 1 | 2 | 4 | |||||||
| 2 | A | 23.12 | 1 | 1 | 2 | ||||||||
| 3 | A | 24.12 | 1 | 2 | 3 | ||||||||
| 4 | B | 24.12 | 2 | 2 | 2 | 1 | 7 | ||||||
| 5 | B | 26.12 | 2 | 3 | 3 | 1 | 9 | ||||||
| 6 | C | 28.12 | 1 | 1 | 1 | 1+ | 4 | ||||||
| 7 | A | 4.01 | 2 | 1 | 1 | 4 | |||||||
| 8 | A | 6.01 | 2 | 1 | 1 | 1 | 1 | 2 | 8 | ||||
|
10 |
A |
15.01 |
1 |
2 |
2 |
1 |
1 |
|
2 |
9 |
|||
| 11 | A | 17.01 | 1 | 1 | 1 | 2 | 5 | ||||||
| (12) | B | 17.01 | 1 | 4 | 1 | 1 | 7 | ||||||
| 13 | B | 18.01 | 2 | 3 | 1 | 2 | 8 | ||||||
| 14 | Ja | 18.01 | 1 | 1 | 1 | 3 | |||||||
| 15 | A | 19.01 | 1 | 1 | |||||||||
| (16) | C | 19.01 | 2 | 1 | 2 | 2 | 7 | ||||||
| 17 | A | 21.01 | 2 | 1 | 3 | 1 | 1 | 8 | |||||
| (18) | A | 23.01 | 3 | 1 | 1 | 1 | 1 | 7 | |||||
| 19 | A | 29.01 | 2 | 1 | 1 | 4 | |||||||
| 20 | C | 2.02 | 1 | 1 | 2 | ||||||||
| (21) | B | 4.02 | 1 | 1 | 2 | 1 | 1 | 1 | 7 | ||||
| 22 | B | 5.02 | 2 | 1 | 2 | 5 | |||||||
| 23 | B | 6.02 | 3 | 2 | 1 | 6 | |||||||
| 25 | B | 13.01 | 1 | 1 | 1 | 3 | |||||||
| 26 | B | 22.01 | 1 | 1 | 2 | ||||||||
| 49 | B | 12-02 | 1 | 1 | 1 | 3 | |||||||
| Total: | 38* 33 | 16 21* | 4* | 6 | 5 | -- | 8 | 2* | |||||
?= Sexo desconocido
* = Un macho adulto y una hembra inmadura escaparon luego de haber sido capturados y transportados a la estación de retención (en Zona A).
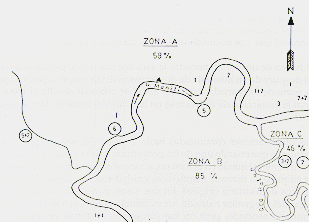
Fig. 4.- División de] área
de estudio en zonas de intensidad de saca.
Los números indican el tamaño de la población remanente
(+ infantes dependientes)
dentro del área de censo (3 km2) después de la saca. Los
círculos indican las manadas intactas.
Zona A: Saca intensiva. Se cosechó el 58% de la población total quedando en el terreno dos manadas multíparas completas; la pareja reproductora con dos crías de una manada multípara; la mitad de una manada multípara (incluyendo la hembra reproductora) y otros tres individuos.
Zona B: Saca muy intensiva. Se cosechó el 85% de la población total, quedando sólo una manada completa y dos individuos adiciona les.
Zona C: Saca semi-intensiva. Se cosechó el 46% de la población, quedando una manada completa y una pareja incipiente.
Efectos de la captura y saca en la población
Debido a su importancia práctica en el manejo del recurso, el estudio sobre los efectos inmediatos de la captura fue orientado hacia los siguientes aspectos: 1) Los efectos de la extracción de individuos en la cohesión social de las manadas remanentes; 2) la propensión relativa de los individuos de diferentes categorías a ser capturados; 3) las consecuencias de la extracción de uno o ambos progenitores en la supervivencia de sus crías no capturadas, y 4) el efecto de la captura y retención de las hembras preñadas en el desarrollo de la gestación y cría. Los resultados obtenidos fueron los siguientes:
1. El hecho de que en 12 manadas la saca fue solamente parcial o completada después de un intervalo de dos semanas desde que se capturó la primera parte de la manada, permitió estudiar con más detalle el efecto que tuvo la extracción de individuos en la cohesión social de la manada residual.
Las observaciones continuadas hasta la 3ra o 5ta semana post-captura, indicaron que cuando uno de los progenitores de la manada afectada (macho o hembra) quedó en el terreno, acompañado por uno o más miembros de su prole, la manada mantuvo su unidad social y continuó viviendo en su área domiciliaria original. En dos casos, tuvimos la impresión que el lugar del progenitor extraído fue eventualmente ocupado por otro adulto errante o asociado, pero no fue constatado en forma definitiva. En los pocos casos donde un individuo quedó en el terreno, éste generalmente no permaneció por mucho tiempo en el área original de su manada capturada, sino divagaba más extensamente en el área circundante, regresando esporádicamente al lugar de origen.
2. En una saca "normal" con fines de aprovechamiento racional, se capturan solamente individuos que demandan una inversión razonable de tiempo y esfuerzo; los individuos "difíciles" se quedan en el terreno. Para que una saca "normal" de Cebuella resulte una extracción equitativa de las diferentes categorías de edad y sexo, consideramos como "cosechados" a aquellos individuos que pudieron ser capturados dentro de los tres primeros días, descartando del análisis los individuos "difíciles". La Fig. 5 indica la proporción de individuos para cada categoría expresada en porcentaje del total de individuos capturados. Los resultados indican que los individuos obtenidos en la saca "normal" se distribuyen equitativamente en las diferentes categorías, salvo los juveniles que parecen más propensos a la captura que los individuos de otras categorías; sin embargo, esta diferencia estadísticamente no es significativa.
La proporción macho/hembra en la muestra
de la saca "normal" fue 46:42, lo que indica que si consideramos
todas las categorías en conjunto, individuos de ambos sexos son capturados
en iguales proporciones. Sin embargo, la Fig. 6 sugiere que entre los subadultos,
los machos serían más propensos a la captura; pero el número
reducido de subadultos en la muestra (7 machos y 7 hembras) no nos permite
dar conclusiones definitivas. En cuanto a los adultos, nuestras observaciones
cualitativas durante las capturas, indicaron que las hembras reproductoras
son marcadamente más propensas a ser capturadas que sus respectivos
machos. La ilustración cuantitativa es que en 86% de los casos las
hembras reproductoras fueron atrapadas en el primer o segundo día;
mientras que en 62% de los casos ocurrió con los machos. A menudo,
éstos fueron atrapados después de 6 a 20 días de la captura
de sus respectivas parejas, y en tres casos los machos no pudieron ser capturados.
Esta diferencia en propensión es particularmente notoria cuando la
manada está con crías dependientes, que son predominantemente
cargadas y cuidadas por el macho, es característico que la hembra sea
una de las primeras en bajar a comer el cebo colocado en la trampa (las hembras
preñadas y lactantes parecen tener un apetito particularmente voraz),
mientras que el macho permanece en lo alto del árbol, cargando o vigilando
a las crías y observando lo que sucede abajo. En una saca "normal",
esta diferencia en comportamiento entre hembras y machos reproductores resultará
una saca desproporcional de hembras lo que, a su vez, afectaría la
recuperación posterior de la población remanente.
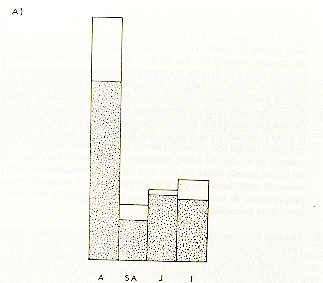
Fig. 8. - Las proporciones de animales
capturados en una saca "normal"
(barras punteadas) y de animales "difíciles" (barras blancas)
en la muestra total capturada.
Fig. 6. - Composición (hipotética)
de la población remanente de
C. pygmaea. (barras sombreadas) después de una saca "normal".
3. En cinco oportunidades capturamos hembras que estaban lactando crías. Las crías no fueron capturadas, sino quedaron en el terreno bajo el cuidado de sus respectivos padres (u otros machos adultos). En dos casos, las crías eran todavía muy pequeñas (menos de un mes de edad) y sus respectivas madres fueron examinadas y liberadas al día siguiente de su captura.
Las observaciones posteriores, que fueron continuadas hasta un mes después de la liberación, indicaron que no hubo abandono o rechazo de las crías, sino ambas madres asumieron normalmente la crianza de su prole. En los tres casos restantes, las crías eran algo mayores (1,5 a 2 meses de edad) y aun cuando todavía recibían leche materna aparentemente no dependían únicamente de ella. En dos de los casos se observó que pese a la ausencia de las hembras, los machos continuaron la crianza de sus respectivas proles. Las crías fueron capturadas tres semanas después de la captura de sus respectivas madres y estaban en aparente buen estado físico. En el tercer caso, el padre y las crías no fueron vistos después de la captura de la hembra (manada 1b); por lo tanto, no se pudo constatar si el macho continuó en la crianza de su prole. Estas observaciones indicarían que la extracción de la hembra no afectaría la supervivencia de sus crías que no dependen únicamente de la leche materna. En los casos donde el padre de las crías fue capturado, la madre continuó la crianza en forma normal.
En una ocasión (manada 17), capturamos ambos progenitores de la manada, quedándose en el terreno un infante dependiente en compañía de un subadulto. Durante la primera semana post-captura, el subadulto fue observado acompañando y ocasionalmente cargando al infante. En la segunda semana, el infante pasó a formar parte de la manada vecina (manada 18) que ocupó el territorio de la manada capturada, y que también tenía una camada de crías.
4. De las siete hembras capturadas en un estado más o menos avanzado de preñez, tres alumbraron en cautiverio y cuatro abortaron en el segundo o tercer día de cautiverio. De las tres nacidas, dos fueron abandonadas por sus progenitores en el primer o segundo día y la tercera después de 10 días de nacida. La captura de hembras en avanzado estado de preñez en Cebuella generalmente resulta una pérdida del 100% de las crías nacidas o por nacer.
RECOMENDACIONES
En el caso de un aprovechamiento racional justificado de C. pygmaea, la saca de poblaciones naturales debería realizarse preferiblemente desde Marzo hasta Abril y de Agosto a Setiembre. Cebuella presenta dos temporadas principales de nacimientos al año y el período de completa dependencia de las crías es de dos a tres meses (Soini, en prensa).
Considerando que en una población natural el número de machos y hembras es equitativo, la saca debe consistir en un igual número de machos y hembras; particularmente, debe cuidarse que la extracción de hembras reproductoras no sea mayor que el de machos reproductores.
Las manadas con crías cargadas o con hembras en un avanzado estado de preñez no deben ser extraídas.
REFERENCIAS
Abbott, D.H. y J.P. Hearn. 1978. Physical,
hormonal and behavioural aspects sexual development in the marmoset monkey,
Callithrix jacchus. 1. &prod. Fert. 53. 155-166.
Hershkovitz, P. 1977. Living New World Monkeys (Platyrrhini). Vol. 1.
Univ. Chicago Press, Chicago. XIV + 1117 pp. and Figs.
Johnston, G.W., S. Dreizen, y B.M. Levy 1970. Dental development in the
cotton ear marmoset (Callithrix jacchus).Amer J.Phys. Anthrop. 33:41-43.
Soini, P. 1982. Ecology and population dynamics of the pygmy marmoset,
Cebuella pygmaea. Folia Primatol. 39: 1-21.
Soini, P. (en prensa). The pygmy marmoset, genus Cebuella. En A.F.Coimbra-Filho
& R.A. Mittermeir (Eds.). Ecology and Behavior of Neotropical. Vol. 2.
Academia Brasileira de Ciencias.
Soini,P. (este volumen). Ecología y dinámica poblacional del pichico
Saguinus fuscicollis. (Callitrichidae, Primates).
Taber, R. 1971. Criteria of sex and age. pp. 325418. En Wildlife Management
Techniques. The Wildlife Society, Washington, D.C.
Anexo 1. - Inventario regional de las fuentes de exudado
del "leoncito" Cebulla pygmaea
El inventario fue realizado en el período 197-5-1979 y abarcó las cuencas de los Ríos Amazonas, Ucayali, Tapiche, Tahuayo, Manití y Orosa.
| Taxón | Tipo de planta | |
| Anacardiaceae | Spodias mombin | árbol |
| Annonaceae | No identificado | árbol |
| Bombacaceae | Ceíba samauma | árbol |
| Burseracoae | No identificado | árbol |
| Celastraceae | Maytenus sp. | árbol |
|
Combretaceae |
Buchenavia sp. |
árbol |
| Terminalia oblonga | árbol | |
| Compositae | No identificado | liana |
| Convolvulaceae | Maripa sp. | liana |
| Dilleniaceac | Doliocarpus sp. (?) | liana |
| Elaeocarpaceae | SIoanea guianensis | árbol |
| Euphorbiaceae | Croton cuneatus | árbol |
| Flacourtiaceae | Laetía sp. (?) | árbol |
| Casearia duckeana | árbol | |
| Gnetaceae | Gnetum sp. | liana |
| Cuttiferae | Vismia sp. | árbol |
| Hippocrateaceae | Cheiloclinium lineatum | liana |
| Cheiloclinium sp. | liana | |
| No identificado | liana | |
| Leguminosae | Acacia riparia | liana |
| Albizzia sp. | árbol | |
| Bauchinia sp. | liana | |
| Campsiandra laurifolia | árbol | |
| Cassia. sp. | árbol | |
| Dióclea sp. (A) | liana | |
| Dioclea sp. (B) | llana | |
| Entada polistachys | liana | |
| Inga ingoides | árbol | |
| Inga marginata | árbol | |
| inga sp. (A) | árbol | |
| Inga sp. (B) | árbol | |
| Inga sp. (C) | árbol | |
| Inga sp. (D) | árbol | |
| Inga sp. (E) | árbol | |
|
Inga sp. (F) |
árbol | |
| Macrolobium acacieafolíum | árbol | |
| Pithecellobium latifolium | árbol | |
| Pithecellobium sp | árbol | |
| Parkia oppositifolia | árbol | |
| Parkia sp. | árbol | |
| Swartzia pendula | árbol | |
| Malpighiaceae | No identificado (A) | liana |
| No identificado (B) | hana | |
| Meliaceae | Carapaguianensis | árbol |
| Cedrela sp | árbol | |
| Trichilia sp. (A) | árbol | |
| Trichilia sp. (B) | árbol | |
| Menispermaceae | Anomospermum sp. | liana |
| Moraceae | Coussapoa sp. | hemiepifita |
| Myrtaceae | Noidenfificado | árbol |
| Polygonaceae | Noidentificado | árbol |
| Rubiaceae | Uncaria sp. | liana |
| Sapindaceae | Pauffinia sp. (?) | liana |
| Sapotaceae (?) | No identificado | árbol |
| Sterculiaceae | Sterculia aff. rugosa | árbol |
| Vochysiaceae | Qualea amoena | árbol |
| Vochysia diversa | árbol | |
| Vochysia lomatophylla | árbol |
LA FAUNA PRIMATOLOGICA EN AREAS DE JENARO HERRERA
Rolando Aquino
![]()
INTRODUCCION
El presente estudio trata sobre evaluación de la situación actual y observaciones sobre el tamaño, asociaciones y comportamiento de la fauna primatológica en áreas correspondientes a Jenaro Herrera, Departamento de Loreto.
Los censos y las observaciones fueron efectuados en su mayoría en las Quebradas de Curiyacu y Tumbo desde 22 Marzo - 15 de Abril, 15 - 21 Mayo y de 26 Mayo - 15 junio 1978.
DESCRIPCIÓN DE LAS ÁREAS DE ESTUDIO
Las áreas de estudio corresponden a las Quebradas Curiyacu, Aucayacu y Tumbo. Las dos primeras se unen para formar el Caño Supay, el cual desemboca en el Río Ucayali, a unos 5 km aguas arriba de la Localidad de Jenaro Herrera. El Tumbo es afluente de la Quebrada Carahuaite, cuyas aguas igualmente vierten al Ucayali cerca a la Localidad de Bagazán. La fisiografía de las áreas estudiadas corresponde a bosques de colina (Malleux 1975), con pequeñas a medianas elevaciones, acentuándose en dirección a las nacientes de las quebradas. Ecológicamente, pertenecen al Bosque húmedo tropical (Tosi 1960).
Los censos no incluyeron los bosques de bajiales de la zona de Jenaro Herrera, que también alberga poblaciones de diferentes especies de primates.
Características de las quebradas
Quebrada Curiyacu, se caracteriza por su curso estrecho. En época de creciente (Enero-Mayo) mantiene un buen caudal, permitiendo la navegación de embarcaciones pequeñas (de 6 a 7 m de longitud). En vaciante (junio-Diciembre), debido al poco caudal y abundancia de palos caídos, prácticamente es imposible la navegación; sin embargo, una buena lluvia durante esta época es suficiente para que la quebrada recupere su caudal hasta el borde de sus riberas. Por tierra, mediante una trocha que nace en la Quebrada Parnayari, próxima al eje de la Carretera Jenaro Herrera- Colonia Angamos, es fácil llegar al lugar denominado Yarinal, la separación entre la quebrada y la carretera en este punto no supera los 10 km.
Quebrada Aucayacu, de lecho más ancho y profundo, con mayor caudal que la anterior. En época de creciente es posible navegar en embarcaciones de 10 m de longitud. En vaciante debido a la merma considerable de sus aguas y abundancia de palos caídos, la navegación se hace difícil, permitiendo llegar únicamente al sector denominado Maquisapay. La quebrada en mención, al igual que la anterior, fue duramente transitada por extractores de shiringa (Hevea brasiliensis), caucho (Ficus sp.), y cazadores furtivos. En la actualidad, aún penetran cazadores con la finalidad de extraer el último de los recursos mencionados.
Quebrada Tumbo, afluente del Carahuaite, la unión de sus aguas se produce aproximadamente a 25 km de la Localidad de Jenaro Herrera, en dirección Sudeste. El curso de esta quebrada es bastante similar al de Curiyacu, lo cual permite navegar solamente en época de creciente y en embarcaciones de 6 m de longitud.
MATERIAL Y MÉTODOS
La evaluación de los primates fue efectuada mediante censos por transectos, método descrito por Southwick y Cadigan (1972) y modificado por Freese (1975), con algunas variantes en cuanto al campo auditivo-visual y tamaño promedio de manada para algunas especies.
Para el censo en el Curiyacu, sectores del Yarinal y primera ramada, aparte de utilizar las trochas existentes con fines de inventario forestal y de extracción de carne de monte, se abrieron previamente trochas por más de 10 km. En el Aucayacu, abarcó los sectores de Camotal, Limón y Colpa Vieja, áreas que poseen trochas en uso por cazadores. Finalmente en el Tumbo, debido a la existencia de trochas poco definidas, el censo comprendió únicamente ambas riberas de la quebrada. En las tres quebradas (Fig.1) los censos fueron ejecutados a pie, a una velocidad promedio de 1,5 km/hora. El grupo de observadores en la mayoría de los casos estuvo integrado por tres personas. En total fueron censados 79,9 km; de ellos, 77,1 km correspondieron al censo diurno y 2,8 km al nocturno, tal como se detalla a continuación:
| Censos: | Curiyacu | Aucayacu | Tumbo | Total |
| Diurno | 27,2 km | 12,6 km | 37,3 km | 77,1 km |
| Nocturno | 2,8 km | ---- | ---- | 2,8 km |
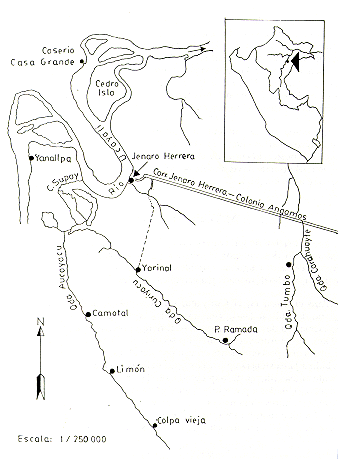
Fig.1. - Cróquis del área de estudio en Jenaro Herrera Dpto. Loreto (Perú)
Adicionalmente, se hicieron observaciones complementarias sobre tamaño de manada, asociaciones y comportamiento de las especies en estudio.
A continuación, se presentan los diferentes campos auditivo-visuales considerados para el análisis de datos.
| Especie | Campo auditivo- visual |
| Saguinus mystax | 60 m |
| Saguinusfuscicollis | 60 m |
| Saimiri sciureus | 100 m |
| Calficebus moloch | 50 m |
| Pithecía monachus | 50 m |
| Cebus albifrons | 150 m |
| Cacajao calvus | 150 m |
| Aotus nancymal | 20 m |
| lagothrix lagothricha | 150 m |
Los campos auditivo-visuales considerados para Cebus albifrons, Cacajao calvus y Lagothrix lagothricha, están basados en las experiencias adquiridas durante las actividades en el bosque.
RESULTADOS Y DISCUSIÓN
Durante el censo efectuado a lo largo de 79,9 km se observaron 104 manadas pertenecientes a nueve especies de primates. El mayor número de encuentros correspondió a Saguinus mystax y Saguinus fuscicollis, con 28 y 26 manadas, respectivamente, seguidos por Pithecia monachus y Callicebus moloch, ambos con 16 y 15 manadas, respectivamente. Cacajao calvus, tras un censo de más de 77 km de longitud, fue localizado en una sola oportunidad lo que permite afirmar que esta especie en el área de estudio, se encuentra en peligro de extinción, si consideramos que hace 15 años atrás los encuentros eran frecuentes (informaciones verbales de moradores antiguos en Jenaro Herrera).
Tamaño de las manadas
Para determinar el tamaño de las manadas de las especies, aparte de considerar las observaciones durante el estudio, también se tomó en cuenta las experiencias adquiridas durante los censos ejecutados en las cuencas del Tahuayo y Blanco, expediciones con similares propósitos a la Estación Biológica Callicebus, en el Río Nanay, Reserva Nacional Pacaya-Samiria y la operación captura de Saguinus mystax, realizada en la cuenca del Manití en 1976. El Cuadro 1 presenta el resumen de las manadas observadas y sus respectivos tamaños, donde podemos notar que los conteos fueron incompletos para la mayoría de las manadas.
S. mystax: Se observó un total de 28 manadas, cuyos conteos mayormente fueron incompletos; sin embargo, en siete de ellos fue posible determinar el tamaño exacto, en un rango de 4 a 8 individuos, lo que permite estimar un tamaño promedio de 6 ind./manada.
S. fuscicollis: De 26 manadas observadas, en siete logramos conteos exactos que fluctuaron desde 3 a 6 individuos, por lo que se deduce como tamaño promedio 6 ind./manada.
S. sciureus: En total se localizaron cuatro manadas y en todos los conteos fueron que dieron cifras mayores de 5 ind./manada. Para el análisis de datos se consideró como tamaño promedio 30 ind./manada.
C. moloch: De 17 manadas localizadas , 15 son consideradas para el censo; los conteos en la mayoría completos, desde 2 a 5 individuos. Por otro lado, datos obtenidos en las cuencas del Manití, Tahuayo, Blanco y Nanay, permiten establecer un tamaño promedio de 4 ind./manada.
P. monachus: Se observaron 20 manadas, cuyos tamaños variaron desde 2 a 4 individuos. Registros similares en otras cuencas y en la Reserva Nacional Pacaya-Samiria, permiten establecer como tamaño promedio 4 ind./manada.
C. albifrons: Se observaron seis manadas, los conteos en todos fueron incompletos. Para el cálculo de densidad, se adoptó como tamaño promedio 10 ind./manada.
C. calvus: Se observó sólo una manada, y en el área del Tumbo. El disparo producido por un cazador dio muerte a un macho adulto, hecho que no permitió determinar el tamaño exacto de manada. No obstante, el conteo incompleto produjo una cifra mayor a 14 ejemplares (incluido el ejemplar muerto), por lo que para el análisis de datos se considera un tamaño de 20 ind./mana-
L. lagothricha: Se localizó un total de seis manadas. En ninguna de ellas fue posible determinar el tamaño exacto. Los conteos incompletos dieron cifras mayores a 10 individuos. Por consiguiente, el tamaño promedio sería 15 ind./manada.
Cuadro 1.- Resumen calendarizado de los censos
de pnmates en áreas de Jenaro 1 lerrera.
| Sector | Long, censada (km) |
Grupos Sm. |
Grupos S.f. |
Grupos S.sc. |
Grupos C.m. |
Grupos P.m. |
Grupos C.a. |
Grupos C.c |
Grupos LI. |
Grupos A.n. |
|
Curiyacu |
30,0 |
>6, >5, |
>4, >4, |
>5 |
4, >3, 3, |
3, >1, 3, |
>4, >5, 8, |
- |
>IO, >4, |
>2, >1, |
| Aucayacu | 12,6 | >6, 7, >5, >3, |
4, >5, >4, |
>120 | 2, 3, | >l, 2, | >9, | -- | -- | -- |
| Tumbo | 37,3 | >3, >5, >4, >3, >3, >2, 6, >2, >4, >4, >3, |
>3, >4, >2, >4, >3 >l 5,>4, >3 ,5, >3, |
>l5, >5 | >2, >l, 3, 2, >l, 4, |
3, 2, >3, 4, 3, >2, |
>4 >7, | >14, | >5, >l0, >7, |
-- |
| Totales | 79,9 | 28 | 26 | 4 | 15 | 16 | 6 | 1 | 6 | 2 |
| Tarnaño promedio |
|
6 | 6 | 30 | 1 | 1 1 | 1 |
| S. m. : | S. mystax | C.a. : | C. albifrons |
| S. f. : | S. fuscicollis | C.c. : | C. calvus |
| S. sc. : | S. sciureus | L. L. : | L. lagothricha |
| C. : | C. Moloch | A. n. : | A. nancymai |
| P. m. : | P. monachus |
A. nancymai: El censo para esta especie fue ejecutado en una sola oportunidad, habiendo localizado dos manadas integradas por más de un ejemplar. Para el cálculo de densidad se consideró 3 ind./manada (Soini y Moya, este volumen).
Densidad poblacional
Los sectores donde se efectuaron los censos fueron considerados como representativos para Jenaro Herrera, puesto que estuvieron sometidos a diferentes intensidades de caza. Las densidades obtenidas por especie se presentan en el Cuadro 2.
S. mystax: Con una densidad general de 6,0 man./km2 y 36,0 ind./km2. Mayor densidad se obtuvo en Curiyacu, mientras que la menor correspondió al Tumbo.
S. fuscicollis: Alcanza una densidad total de 5,6 man./km2 y 33,7 ind./km2. Las densidades parciales en las áreas del Curiyacu, Aucayacu y Tumbo están indicadas en el Cuadro 2.
Ambas especies de Saguinus son bastante comunes en las áreas de estudio e incluso en las cercanías de la Localidad de Jenaro Herrera.
S. sciureus: Con una densidad total de 0,5 man./km2 y 15,6 ind./km2. La mayor densidad fue registrada en Aucayacu.
C. moloch: Con una densidad general de 3,9 man./km2 y 15,4 ind./km2; correspondiendo al Curiyacu la mayor densidad, mientras que la menor fue registrada en Aucayacu.
P. monachus: Su densidad general fue de 4,1 man./km2 0 15,6 ind./km2, obteniéndose mayor densidad en el Curiyacu.
C. albifrons: Alcanza una densidad general de 0,5 man./km2 o 5,1 ind./km2 obteniéndose mayor densidad en el Curiyacu.
C. calvus: Tuvo una densidad general de 0,1 man./km2 0 1,6 ind./km2, constituyendo así la densidad más baja registrada durante los estudios realizados.
L. lagothricha: Se registró una densidad total de 0,5 man./km2 o 7,8 ind./km2. Mayor densidad correspondió al Curiyacu. En Aucayacu esta especie no fue localizada.
Cuadro 2.- Densidades estimadas en las áreas
de estudio bajo la modalidad del censo por transeM Especies a Curiyacu Aucayacu
1 Tumbo Densidad General
|
Especies |
Curiyacu |
Aucayacu |
Tumbo |
Densidad General | ||||
|
Grupo/km2 |
Indiv./km2 |
Grupo/km2 |
Indiv./km2 |
Grupo/km2 |
Indiv./km2 |
Grupo/ km2 |
Indiv./km2 |
|
|
S. mystax |
7,8 | 47,8 | 5,5 | 32,2 | 4,9 | 30,0 | 6,0 | 36,0 |
|
S. fuscicollis |
7,4 | 44,2 | 4,0 | 24,0 | 4,9 | 30,0 | 5,6 | 33,7 |
|
S. sciureus |
0,4 | 11,1 | 0,8 | 24,0 | 0,5 | 16,0 | 0,5 | 15,6 |
|
C. moloch |
5,1 | 20,6 | 3,2 | 12,7 | 3,2 | 12,9 | 3,9 | 15,4 |
|
P. monachus |
5,9 | 23,5 | 3,2 | 12,7 | 3,2 | 12,9 | 4,1 | 15,6 |
|
C. albifrons |
0,7 | 7,3 | 0,5 | 5,3 | 0,3 | 3,5 | 0,5 | 5,1 |
|
C. calvus |
- | - | -- | -- | 0,2 | 3,4 | 0,1 | 1,6 |
|
L. lagothricha |
0,7 | 11,0 | -- | -- | 0,5 | 8,0 | 0,5 | 718 |
Las tres últimas especies no es posible observarlas en las cercanías de la localidad. Las manadas ubicadas correspondieron a los sectores de la primera manada, Camotal, y Tumbo, distantes unos 20, 35 y 30 km, respectivamente, de la Localidad de Jenaro Herrera.
A. nancymai: El censo fue ejecutado en el sector de Yarinal en el Curiyacu. La densidad obtenida fue de 35,7 man./km2 o 107,1 ind./km2. La elevada densidad obtenida indudablemente sobreestima a la verdadera.
Asociaciones interespecíficas
S. mystax: Por lo general se encuentra asociado con Saguinus fuscicollis que ocupa el estrato más bajo del bosque. También con frecuencia los observamos solos, éstos después desmovilizares conjuntamente un buen trecho, a menudo tienden a separarse.
S. fuscicollis: Conjuntamente con S. mystax fueron observados en asociación con C. moloch cuando se hallaban cruzando un aguajal. La manada mixta estuvo conformada por seis individuos de S. fuscicollis, ocho de S. mystax y cinco de C. moloch. Durante la locomoción los S. fuscicollis servían de guía a las otras especies. Aparte, S. mystax y S. fuscicollis se hallaron en asociación con P. monachus hasta en cuatro oportunidades (tres de ellas durante el censo). En los puntos de contacto se notó que P. monachus acostumbra a seguirlos a los dos primeros ocupando el mismo estrato que S. mystax.
Finalmente, C. albifrons fue observado en asociación con L. lagothricha y S. sciureus. En ambos casos, C. albifrons servía de guía a las demás especies. Los Cuadros 1 y 2 muestran las especies halladas en asociaciones.
Observaciones sobre el comportamiento de algunas especies
S. mystax: Normalmente de movilización lenta y silenciosa. En lugares donde son perseguidos por el hombre, al advertir la presencia de éste, emiten vocalizaciones de alarma y huyen mediante saltos cortos, seguidos de movimientos rápidos hasta perderse en la espesura del bosque. Esta reacción naturalmente no sucede en ciertos lugares que no están sujetos a alta presión de caza, corno Curiyacu y Tumbo, donde la presencia humana únicamente causó alarma y en la mayoría de las veces subieron a las copas de los árboles, permaneciendo allí hasta que lograron calmarse para iniciar la retirada sin precipitarse; otras veces permanecieron indiferentes al observador.
S. mystax: Por lo común ocupa el estrato comprendido entre los 4 y 10 m; sin embargo, en varias ocasiones se hallaron a niveles inferiores como el sotobosque; tal es así que en una oportunidad se localizó una manada dispersa, algunos de ellos se hallaban en bejucos bajos, próximos al suelo, en típica posición de descanso, mientras que otros se encontraban en la ribera de la quebrada cazando insectos a no más de 1m de alto. Por otro lado, a sólo 1 km de la Localidad de Jenaro Herrera, hubo una manada, cuya área domiciliaria fue seccionada por la Carretera J. Herrera-Colonia Angamos mediante un despeje de más de 30 m de ancho; en este sector, periódicamente se observaron a los componentes de la manada cruzando la carretera por el suelo, lo que indicaría que éstos son propensos a la actividad terrestre en algunas circunstancias.
S. fuscicollis: Ocupa el estrato más bajo del bosque hasta aproximadamente 6m de alto. Igual a la anterior, esta especie reacciona ante la presencia humana mediante vocalizaciones de alarma, y acostumbra movilizarse en posición vertical en los troncos y ramas de los árboles. No huyen mientras no hayan detectado al observador, para lo cual incluso tienden a acercarse mediante saltos cortos pero rápidos. Tan pronto advierten la presencia humana, huyen rápidamente. También se ha comprobado que S. fuscicollis, después de varios minutos de alerta, termina por ser indiferente al observador y reinicia sus actividades normalmente. En una oportunidad, en la Estación Biológica Callicebus - Río Nanay, durante 4:40 h logré seguir una manada conformada por 8 individuos, quienes inicialmente se mostraron alarmados e iniciaron la huida rápida; después de casi una hora de seguimiento, éstos paulatinamente fueron perdiendo el temor, hasta llegar a ser indiferentes a mi presencia.
S. fuscicollis, por lo general se movilizan entre 1 a 6 m de alto, para lo cual incluso aprovechan palos caídos, y se dispersan en más de 50m de ancho, pero mantienen la dirección de movilización mediante vocalizaciones de larga distancia. Son mucho más activos en horas matutinas y al atardecer, por lo que frecuentemente los contactos se producen cuando se hallan movilizándose o en plena alimentación. A mediodía normalmente descansan y permanecen silenciosos.
C moloch: Especie de movimiento lento y casi silencioso, acostumbra caminar por las ramas y sólo realizan saltos cuando la separación de las ramas o árboles supera a sus miembros extendidos. Ante la presencia del hombre reacciona emitiendo localizaciones de alarma de tono seco y corto, huyen rápidamente o se ocultan entre los follajes o bejucos; algunas veces incluso tienden a lanzarse a tierra y corren en busca de huecos o cualquier otro medio de protección.
P. monachus: Normalmente son de movimiento lento ; realizan saltos suaves y casi silenciosos. Acostumbra caminar entre las ramas y muchas veces permanecen sentados por varios minutos observando sus alrededores: En lugares de alta presión de caza, tan pronto advierten la presencia humana huyen velozmente mediante saltos que superan los 10 m de largo o simplemente se ocultan en las copas más frondosas de los árboles. Sin embargo, durante el presente estudio, principalmente en las Quebradas Tumbo y Curiyacu, reaccionaron a nuestra presencia con vocalización de alarma y erizando los pelos, peculiaridades que fueron acompañadas de movimientos laterales y rápidos del cuerpo; otras veces hicieron ademanes de bajarse de los árboles emprendiendo luego la huida mediante saltos prodigiosos.
C. albifrons: Especie de movimiento rápido y salto largo, cuya movilización ruidosa puede ser percibida a más de 100 m de distancia. En una oportunidad, C. albifrons fue localizado alimentándose de frutos de shebón (Scheelea sp.), pero tan pronto descubrieron nuestra presencia, emprendieron la fuga dispersándose totalmente la manada. Minutos más tarde, en un acto probablemente de reagrupamiento, escuchamos vocalizaciones de larga distancia. C. albifrons es una especie que, cuando uno imita su vocalización tiende a contestar y aproximarse, oportunidad que aprovechan los cazadores para cazarlo.
C. calvus: Monos bulliciosos, cuyas vocalizaciones son tan peculiares que los diferencian de los demás, por lo que fácilmente pueden ser identificados a distancias mayores de 100 m. Precisamente a raíz de esta peculiaridad, la única manada fue localizada en un "aguajal" cuando estaban alimentándose de frutos de aguaje (Mauritia flexuosa). Lamentablemente, la detonación de un arma de fuego produjo desconcierto en la manada, huyendo desesperadamente y guiados por los machos adultos. Conjuntamente con la manada se observó decenas de rapaces, volando detrás del grupo, entre ellas destacaba la presencia del águila monera Harpia harpyja.
L. lagothricha: Especie que gusta caminar por las ramas y realizar saltos largos, cuando la separación entre dos árboles es amplia, ocasionando ruidos que pueden escucharse a más de 100 m. Ante la presencia del hombre huyen rápidamente, emitiendo vocalizaciones de alarma en tono grueso y grave, mientras que algunos acostumbran sacudir fuertemente las ramas.
Evaluación de la situación actual de las especies en Jenaro Herrera
C. rygmaea, S. mystax, S. fuscicollis, S. sciureus y A. nancymai son frecuentemente observadas, incluso en las cercanías de la Localidad de Jenaro Herrera. La caza de estos animales para el consumo humano es esporádica, por lo que se los puede considerar comunes.
C. moloch y P. monachus: En la actualidad son eventualmente cazados para el consumo de subsistencia, siendo rara su presencia en las cercanías de la localidad de Jenaro Herrera. En áreas del Curiyacu y Tumbo, son los únicos lugares donde se les puede observar con cierta frecuencia. En consecuencia, ambos se encuentran en situación vulnerable.
C. albifrons y L. lagotricha: Son posibles de observarlos en lugares distantes, es decir a un día de camino en dirección oriental desde la Localidad de Jenaro Herrera. Actualmente, ambos están sometidos a la más alta presión de caza, principalmente a lo largo del eje de la Carretera Jenaro Herrera-Colonia Angamos. En consecuencia los consideramos en situación amenazada
C calvus- Se trata de la única especie que no fue posible encontrar con regularidad. El motivo principal es indudablemente la fuerte presión de caza que viene soportando, muchas veces sin otro objetivo que el de obtener la cría para ser vendida como mascota. En consecuencia a esta especie podemos considerarla en vías de extinción para esta parte del oriente peruano.
Finalmente, Ateles paniscus y Cebus apella fueron encontradas en calidad de mascotas en la Localidad de Jenaro Herrera; sin embargo, no fueron observadas en el terreno, por lo que deducimos que han sido completamente o casi exterminadas en el área de estudio.
CONCLUSIONES
Durante los censos realizados en las Quebradas de Curiyacu, Aucayacu y Tumbo se registraron Cebuella pygmaea, Saguinus mystax, Saguinus fuscicollis, Saimiri sciureus, Callicebus moloch, Pithecia monachus, Cebus albifrons, Aotus nancymai, Cacajao calvus y Lagothrix lagothricha. Además, Ateles paniscus y Cebus apella fueron hallados como mascotas en la misma localidad.
Las especies más comunes son: Cebuella pygmaea (no considerada en el censo), S. mystax y S. fuscicollis, alcanzando las dos últimas las más altas densidades. C. moloch y P. monachus también alcanzaron densidades relativamente altas, principalmente en la Quebrada Curiyacu.
En S. sciureus y C. calvus, encontramos las más bajas densidades. En la primera, la explicación entre otros factores podría ser que éstos tienen como hábitat preferencial a los bajiales y no las colinas altas.
La mayor incidencia de caza vienen sufriendo C. calvus, L. lagothricha y C. albifrons, La apertura de la Carretera Jenaro Herrera - Colonia Angamos, influye en la elevada presión de caza, pues a través de ella los cazadores llegan a sectores mucho más alejados, poniendo incluso en grave peligro de extinción a C. calvus y A. paniscus.
REFERENCIAS
Freese, C. 1975. Informe final sobre censo de primates en el Perú. OPS., Wash., D.C. 37pp.
Malleux, J. 1975. Mapa Forestal del Perú. (Memoria Explicativa). Universidad Nacional Agraria La Molina, Lima. 161 pp.
Soini, P . y L. Moya. 1987. Informe sobre los resultados de los estudios primatológicos en la zona norte de la Isla Iquitos. Ministerio de Agricultura, R.A. XXII -Iquitos.
Southwick, C. y F. Cadigan. 1972. Population studies of Malaysian primates. Primates 13:1 -18.
Tosí, J. 1960. Zonas de vida natural en el Perú.
Inst. Inter. de Ciencias Agric. Bol. Téc. 5.211pp.

ASPECTOS DE LA ADAPTACION DE Saimiri boliviensis peruviensis y S. sciureus macrodon A NUEVAS FUENTES ALIMENTICIAS EN LA AMAZONIA PERUANA
Filomeno Encarnación
![]()
INTRODUCCION
El presente trabajo es el resultado de los análisis de comportamiento de Saimiri boliviensis peruviensis y S. sciureus macrodon, los que progresivamente se adaptan al consumo temporal de frutos inmaduros (choclos) de maíz (Zea mays) y de otros frutales cultivados, inducidos por una serie de factores ambientales y biológicos. Las observaciones y muestreos realizados de las manadas de frailes se efectuaron de Junio a Diciembre 1977, durante los programas experimentales de manejo extensivo y en cautiverio de primates no humanos.
Las áreas de estudio y observación están localizadas en las márgenes de los Ríos Putumayo, Tigre, Marañón, Ucayali, Amazonas y algunos afluentes menores, los que han estado sometidos a intensivas actividades agrícolas periódicas o cíclicas (cultivos de roza, tala y quema) desde uno o más años. Los cultivos son principalmente el maíz, yuca (Manihot esculenta), arroz (Oriza sativa), frijoles (Phaseolus sp.), chiclayos (Vigna sinensis).
Entre los frutales preferidos por los frailes en su dieta diaria se hallan Musa sapiens, M. paradisiaca (plátanos diversos), caimito (Pouteria caimito, P. guyanensis), guava (Inga edulis), shimbillo (Inga sp.), uvilla (Pourouma cecropiaefolia).
MATERIALES Y MÉTODOS
Materiales
Fueron observadas y estudiadas 36 manadas de frailes (16 del tipo gótico y 20 del tipo romano), así como otras especies animales que frecuentan los terrenos cultivados y se refugian en los bosques adyacentes. Para las observaciones complementarias, se emplearon racimos maduros de plátanos de la variedad "inguiri" y mazorcas inmaduras de maíz (choclos) de las variedades "polvo sara" o "shishaco". Se atraparon 220 ejemplares (81 del tipo gótico y 139 del tipo romano), pertenecientes a 12 manadas; en éstos se efectuaron observaciones experimentales sobre aceptación, palatabilidad y consumo de seis frutos diferentes durante los 10 primeros días en cautiverio.
Métodos
Se empleó la siguiente metodología:
1. Localización de los territorios y hábitats (área de actividad), siguiendo los transectos y senderos utilizados por los pobladores ribereños.
2. Las observaciones de comportamiento y hábitos alimenticios se realizaron en el bosque y los cultivos, mediante el uso de binoculares, entre las 06:00 a 09:00 y de 16:00 a 18:00 h (hora local), totalizando 498 horas en 166 repeticiones matutinas, y 286 horas en 142 repeticiones vespertinas. No todas las observaciones fueron en días sucesivos; sin embargo, se procuró obtener el ritmo de tres horas de observación por las tardes, prolongándose este tiempo de acuerdo a las circunstancias. Durante este lapso, en el área despejada y chacras, la humedad relativa es alta y la radiación solares baja (Thorington 1968).
3. Colocación de señuelos o cebos (plátanos) en el área de actividad de las manadas y observación de la conducta frente a los mismos, verificando la aceptación o no de tales señuelos.
4. Atrapamiento, mediante el empleo de trampas, con fines de muestreo, aprovechamiento racional y otros estudios.
5. Observación de la aceptación y consumo de choclos, plátanos, caimitos, guava, shimbillo y uvillas, durante los primeros días de cautiverio en el local cuarentenario del Centro de Reproducción y Conservación de Primates No Humanos de Iquitos.
6. Encuesta entre los pobladores y agricultores de la zona, sobre la identificación de los invasores de cultivo y los daños ocasionados por éstos.
DESCRIPCIÓN DE LAS ÁREAS DE ESTUDIO
Los muestreos (atrapamiento) y observaciones se ejecutaron en bosques de la llanura meándrica o "várzea estacional" (con inundación estacional), y en las colinas o "tierra firme" (Cain et al. 1956, Prance 1979). Estos terrenos son adyacentes a los Ríos Putumayo, Tigre, Marañón, Ucayali y Amazonas; corresponden a la clasificación forestal del Perú de Malleux (1975) a bosques aluviales clases 11 y 111, y bosques de colina clase 1. Según ONERN (1976), pertenecen a la zona de vida de bosque tropical y transición a bosque húmedo tropical. La inundación periódica por aguas "blancas" Y "claras" (Sioli 1974) durante la estación lluviosa o "creciente" (Enero-Abril), y la consecuente sedimentación y deposición aluvial en la llamada "várzea estacional" (Marlier 1979, Prance 1979), o tahuampa (Encarnación 1985), especialmente en las orillas y terrenos adyacentes a los grandes afluentes del Amazonas, ejercen efectos fertilizantes y renovables, mejorando el rendimiento agrícola de los suelos durante la estación seca o "vaciante" (Mayo-Diciembre), determinando la acción humana de tala-roza para destinarlo al cultivo y cosechas efímeras (tres meses), tales como arroz, maíz, frijol, chiclayo, yuca y otros. En las partes altas o "restingas" inundables en "crecientes" extremas, así como en colinas se cultivan maíz, yuca, y frutales como plátanos, caimitos, uvillas, guava, shimbillos, etc.
En todos los tipos de bosques se siguen las fases de la agricultura errante tradicional (Meggers 1973), mediante la tala-roza-quema y sembrado.
Los terrenos estudiados con cultivos de maíz y plátanos tenían de uno a tres años o más de antigüedad (Cuadros 1 y 2 ) considerando la rotación anual.
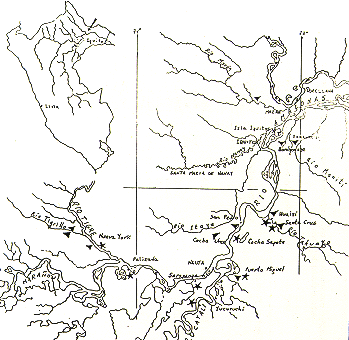
En la Fig. 1 y el Cuadro 1,,;e indican las localidades estudiadas y algunas notas acerca de la actividad agrícola en ellas; estas localidades fueron:
- Nueva York (R. Tigre), margen izquierda del Marañón.
- Palizada, margen derecha del Marañón.
- Sarapanga y Yucuruchi, confluencia de los Ríos Marañón y Ucayali.
- Puerto Miguel (Quebrada Yarapa), margen derecha del Amazonas y del Ucayali.
- Nazareth, margen derecha del Amazonas.
- Cocha Sapote (frente a San Joaquín de Omagua), margen derecha del Amazonas.
- San Pedro, margen izquierda del Amazonas.
- Huaisi y Santa Cruz (R. Tahuayo),margen derecha del Amazonas.
- Bombonaje (Quebradas Yanayacu y Bombonaje), margen derecha del Amazonas.
- Bellavista y San Martín (R.Yuvineto), margen derecha del Putumayo.
Cuadro 1.- Localidades de estudio de Saimiri spp. (frailes) y sus relaciones con la actividad agrícola de Zea mays (maíz) en terreno de Tahuampa estacional.
| Localidades de estudio | Tipo ambientes | Relación con la agricultura | Tipo de Samiri spp. | ||||||
|
B. |
B. |
B. Colina |
Grado de actividad |
Consumidores |
Edad de cultivos |
Gótico | Romano | ||
|
Clase II |
Clase II |
Clase III |
1 a 3 | más de 3 | |||||
|
Nueva York (Río Tigre) |
- |
+ |
- |
x |
- |
+ |
- | + | - |
|
Palizada (Río Marañón) |
- |
+ |
- |
xx |
- |
+ |
- | - | + |
|
Sarapanga (Ríos Ucayali y Marañón) |
- |
+ |
- |
xxxx |
+ |
+ |
- | + | |
|
Yucuruchi (Ríos Ucayali y Marañón) |
- |
+ |
- |
xxxx |
+ |
+ |
- | + | |
|
Puerto Miguel (Quebrada Yarapa) |
- |
+ |
- |
xxx |
+ |
+ |
- | + |
+ |
|
Nazareth (Río Amazonas) |
- |
+ |
- |
xxx |
- |
+ |
- | - | + |
|
Cocha Sapote (Río Amazonas) |
- |
+ |
- |
xxx |
- |
+ |
- | - | + |
|
Cocha Urcu (Río Amazonas) |
- | - |
+ |
xx |
- |
+ |
- | + | - |
|
San Pedro (Río Amazonas) |
- | - |
+ |
xx |
- |
+ |
- | + | - |
|
Huaisi (Río Tahuayo) |
- |
+ |
- |
xxxx |
+ |
+ |
+ |
+ |
|
|
Santa Cruz (Río Tahuayo) |
- | - |
+ |
xxx |
- |
+ |
- |
+ |
- |
|
Bombonaje (Quebradas Bellavista y San Martín |
+ |
- |
+ |
xx |
+ |
- |
+ |
- | |
|
(Río Yuvincto) |
+ |
+ |
+ |
x |
- |
+ |
- | + |
- |
|
Isla Iquitos (Río Amazonas) |
- |
+ |
- |
xxxxa |
+ |
- |
+ |
+b |
+b |
Cuadro 2.- Factores que inducen al aprendizaje y consumo de maíz
y registro de observaciones del comportamiento en ocho manadas de frailes.
|
Localidades |
Grado de actividad agrícola |
TC años |
AOC |
Repeticiones de las observaciones |
|||||||
| 1 a 3 | más de 3 | Diurno | Nocturno |
1977 |
1978 |
||||||
| A.M. | P.M. | G.C. | A.M. | P.M. | G.C. | ||||||
| Sarapanga | xxxx | - | + | DD | NN | 6(18) | 2(6) | 11(30) | 9(20) | ||
|
Yucuruchi |
xxxx |
- |
+ |
D |
N |
n.o. |
n.o. |
8(24) |
7(13) |
n.o. |
|
| Puerto Miguel | xxx | + | DD | N | n.o. | n.o. | 8(24) | 5(10) | n.o. | ||
| Nazareth | xxx | + | - | DDD | N | 12(36) | 8(19) | a | 21(65) | 16(32) | a.d. |
| Cocha Sapote A | xxx | + | - | DD | N | 20(63) | 23(45) | - | 17(51) | 15(30) | a.d. |
| Cocha Sapote B | xxx | + | - | DD | N | 18(54) | 25(52) | a | 12(36) | 9(18) | a.d. |
| Huaisi | xxxx | - | + | DD | NN | 15(45) | 7(14) | +++ | 10(27) | 5(12) | +++ |
| Santa Cruz | xxx | + | D | NN | 2(5) | 3(5) | - | 6(20) | 8(10) | - | |
|
TOTAL |
|
|
73(221) 68(141) |
93(277) 74(145) |
|||||||
| . |
Los números fuera de paréntesis indican repeticiones, y los incluidos en paréntesis indican el total horas de bservaciones |
CC |
grado de consumo |
|
A.M. |
Observaciones en horas de la mafiana. |
a |
aprendizaje (o aceptación) |
|
P.M. |
Observaciones en horas de la tarde. |
d |
Deshojado de nuevas mazorcas |
|
AyB |
Areas distantes entre 5 a 8 km en la misma localidad |
TC |
Tiempo transcurrido desde que se inició el cultivo de maíz |
|
n.o. |
No observado | ||
|
AOC |
Acción de otros consumidores |
NOTA: Las observaciones corresponden de 06:00
a 09:00 y de 15:00a 18:001% (hora local).
Localidades de Estudio |
|
|
RESULTADOS Y DISCUSION
La deforestación (o despeje) y quema de la vegetación de los terrenos para los sembríos de maíz y otros cultivos, disminuyen las áreas boscosas naturales donde habitan los animales silvestres arborícolas y terrestres, induciéndolos a ciertos cambios en sus patrones de búsqueda y obtención de fuentes de alimentos en forma directa o indirecta, afectando los cultivos del hombre por acciones sinergéticas de consumo.
El Cuadro 3 y Fig. 2, muestran las preferencias de los frailes, de diferentes localidades por las frutas cultivadas y endémicas de la amazonía, además se aprecian las relaciones entre el grado de aceptación de los choclos, por los frailes, y la amplitud e intensidad de este cultivo.
En el Cuadro 3, se aprecia que después de transcurridos cinco días del atrape, todos los grupos de frailes consumieron los choclos y plátanos con intensidad similar. Por consiguiente, se establece una relación directa entre el tiempo transcurrido desde los primeros despejes con fines agrícolas y de adaptación al consumo de los choclos mediante el aprendizaje progresivo del deshojado de nuevas mazorcas (Rumbaugh 1968, Kurnmer 1973).
Se supone que estos cambios en los patrones de búsqueda y obtención de alimentos, pueden ser explicados de la manera siguiente:
1. Origen y procedencia de las especies cultivadas consumidas:
El maíz y el plátano son plantas exóticas a la agricultura de los ambientes de la amazonía, que introducidas , naturalizadas Y cultivadas en gran escala para el consumo humano, se presentan como nuevas fuentes de alimentos en los terrenos contiguos a los bosques o ambientes naturales de los animales silvestres. Por el contrario, los frutales (Cuadros 4 y 5) cultivados de caimito, guavas, shimbillo, uvilla, etc., tienen aspecto y sabor similares a sus congéneres y especies silvestres, con fructificación periódica anual y baja productividad.
2. Inducción por otros consumidores:
En los terrenos adyacentes a Puerto Miguel, Nazareth, Cocha Sapote (A* y B) (*Se designan con estas letras, los lotes de terrenos distantes de 5 a 8 km en la misma localidad) , Huaisi y Santa Cruz, se efectuaron observaciones comparando la aceptación progresiva al consumo de choclos y el deshojado de las mazorcas (Cuadros 2 y 3).
Cuadro 3.- Registro de muestreo (atrapamientos)
de Saimiri spp. y su comportamiento ante los frutos cultivados.
|
Localidades |
Muestreos |
Consumo de frutos (campo) |
Aceptación (cautiverio) |
Tipos |
|||||||
| Tamaño manada estimado | individuos atrapados |
plátano |
maíz |
caimito | guava | shimbillo |
uvilla |
plátano |
maíz |
||
| Sarapanga | 35 | 29 | + | + | + | + | + | + | i | i |
R |
| Yucuruchi | 60 | 39 | + | + | + | + | + | + | i | i |
0 |
| Palizada 9 | 30 | 7 | - | - | no | no | no | no | 1 | 4 |
M |
| Cocha Sapote A | 60 | 7 | + | - | + | + | + | + | i | 5 |
A |
| Cocha Sapote B | 90 | 25 | + | - | + | + | + | + | i | 2 |
N |
| Cocha Sapote C | 40 | 18 | + | - | + | + | + | no | i | 2 |
O |
| Huaisi | ~60 | 14 | + | + | + | + | + | + | i | i | |
| Puerto Miguel | 40 | 11 | + | - | + | + | + | + | i | 5 |
G |
| Cocha Urcu, A | 40 | 35 | - | - | + | + | + | + | 1 | 5 |
O |
| Cocha Urcu B | 30 | 11 | - | - | + | + | + | + | 1 | 3 |
T |
| San Pedro A | 935 | 7 | + | - | + | + | + | + | i | 3 |
I |
| San Pedro B | 35 | 17 | + | + | + | + | + | i | 3 |
C |
|
| O | |||||||||||
| 1,2, .. | : | días transcurridos para la aceptación |
|
i |
: |
aceptación inmediata |
|
no |
: |
no observado |

Fig. 2. - Comportamiento en cautiverio
de los Saimiri spp. ante los frutos cultivados.
Se deduce que estos fenómenos están relacionados
y son el resultado de continuas visitas a las "chacras" (con plantaciones
de maíz) , donde la fauna entomológica es abundante y se hallan
inducidos por visualización directa de alimentos consumidos y restos
dejados por mamíferos y aves frugívoros (Cuadro 6 ). Estos últimos
son consumidores naturales oportunistas, invasores de cultivos de maíz
y plátano, que en sus incursiones diurnas y nocturnas dejan abiertas
las mazorcas inmaduras de maíz o choclos y los plátanos, exponiéndolos
al consumo por los monos frailes.
Cuadro 4.- Plantas incluidas en la alimentación
o sustrato de alimentos de Saimiri boliviensis peruviensis
Localidades: Huasai, Nazareth y Sarapanga
| Especies | Nombre vulgar | E | F | M | A | M | J | J | A | S | O | N | D |
| Nativas | t | ||||||||||||
| Clarisia sp. | capinurí | f | f | f | |||||||||
| I%,a sp. | shImbillo (restiriga) | f | f | f | |||||||||
| Parkia igneiflora | gomaguayo | F | F | F | F | ||||||||
| RJiee(lia sp. | charichuela (chica) | f | f | f | |||||||||
| Rheed la sp. | charichuela(verrucosá | f | f | f | |||||||||
| Inga sp. | shimbillo (orillas y cochas) | f | f | f | f | ||||||||
| Heliconia sp. | platanillo, pico de loro | Fi | Fi | Fi | Fi | ||||||||
| Physalis angulata | mullaca, | f | f | f | f | f | |||||||
| Cultivadas | |||||||||||||
| Zea niays | maíz | f | f | f | f | f | f | ||||||
| Pourouma | uvilla |
f |
f |
f |
f |
f |
|||||||
|
cecropiafolia |
|||||||||||||
|
Inga sp. |
tablacho shimbillo |
f |
f |
f |
f |
||||||||
|
Musa sp. |
plátano guineo |
|
f |
f |
f |
f |
|||||||
|
Pouteria caimíto |
caimito | f | f | f | f | ||||||||
|
Inga edulis |
guava |
f |
f |
f |
f |
||||||||
|
f |
f |
f |
f |
f |
|
F. : |
cazan los insectos que polinizan esta especie |
|||
|
Fi : |
cazan las arañas que se guarecen entre las brácteas |
|||
|
f : |
frutos |
|||
Cuadro 5.- Plantas incluidas en la alimentación
o sustrato de alimentos de Saimiri ociurrus macrodón "fraile
Localidades -Cocha Urcu, Santa Cruz y Bombonaje.
| Especies | Nombre vulgar |
E |
F |
M |
A |
M |
J |
J | A | S | O | N | D |
| Nativas |
|
||||||||||||
| Parkia oppositifolia |
gomaguayo |
F |
F |
F | F | ||||||||
| Strichnos sp. |
(frutos dulces) |
f |
f | f | |||||||||
| Naucleopsis sp. | tamamuri | f | f | ||||||||||
| Chrisophillum sp. | caimitillo | f | f | f | |||||||||
| Duroia hirsuta | sachacaimito | f | f | f | |||||||||
| Duroia longifolia | sachacaimito |
|
f | f | f | ||||||||
| Cultivadas |
|
|
|||||||||||
| Musa sp. |
plátano guineo |
f |
f |
f |
|||||||||
| Pourouma cecropiaefolia |
uvilla |
f |
|
|
f |
f |
f |
||||||
| Pouteria guyanensis |
caimito |
f |
f |
f |
f |
||||||||
| Pouteria caimito |
caimito |
f |
f |
f |
f |
||||||||
| Inga edulis |
guava |
f |
f |
f |
|
F |
: |
cazan los insectos que polinizan la especie |
||
|
f |
: |
frutos |
||
3. Disponibilidad de alimentos:
Los frailes, en su actividad diaria de búsqueda de alimentos, principalmente insectos (Thorington 1968; Soini y Moya, este volumen), incursionan en las chacras con maíz. En la Fig. 3 se indican las preferencias alimenticias de los frailes según las disponibilidades, con la secuencia de prioridad en las chacras de uno a más años: 1) Insectos (Orthoptera), 2) larvas de insectos, 3) arácnidos, 4) frutos de Physalis, y 5) choclos. Luego de tres años se advierte alteraciones en el orden de las prioridades: 1) choclos, 2) insectos (Orthopera), 3) larvas de insectos, 4) arácnidos, y 5) frutos de Physalis.
Estos fenómenos fueron comprobados en ocho manadas que habitan las localidades mencionadas en el Cuadro 2, encontrándose que después de 158 h de observaciones, en 52 eventos matutinos y 121 h en 59 eventos vespertinos durante 1977, que en los "maizales" de Nazareth y Cocha Sapote B, el consumo de choclos por los frailes era sinergia complementaria iniciada por las aves y los mamíferos, con ocurrencia del deshojado de nuevas mazorcas. Por comparación, en el lote de Sapote A y en Santa Cruz, no se observó aceptación de los choclos. En 1978, las observaciones fueron más minuciosas, con 172 h en 53 eventos matutinos, y 90 h en 48 eventos vespertinos, observándose que en los terrenos de Nazareth y Cocha Sapote B, habían preferencias e intenso deshojado de las mazorcas como resultado del progresivo aprendizaje (Wilson 1975), mientras que en Santa Cruz y Cocha Sapote A, no se apreciaron cambios significativos respecto a la aceptación y aprendizaje del deshojado de mazorcas.
Cuadro 6.- Consumidores de Zea mays (maíz) y Musa sp. (plátanos) que inducen similar comportamiento en los frailes Saimiri sciureus.
| Actividad Nocturna | Especie | Nombre vulgar | Clase |
| Philander opposum | Zorro Plomo | mamifero | |
|
Diurna |
Brotogeris cyanoptera | pibuicho cachete amarillo | ave |
| 8. sanctithonw | pihuicho frente am^ | ave | |
| 8. versicolorus | pihuicho ala blanca | ave | |
| Amawnaspp. | loros | ave | |
| Pionites nwlanocephala | chirriclé | ave | |
| Cacicus cela | paucarillo | ave | |
| Psarocoflus angus t lfrons | bocholocho | ave |
4. Adaptación al consumo de cultivo en las áreas estudiadas:
Las manadas de frailes del tipo "romano", que habitan los bosques adyacentes a Sarapanga, Yucuruchi y Huaisi, consumen ávidamente los choclos y plátanos "in situ". Estos cultivos periódicos, en amplias extensiones, se repiten por más de 10 años.
Observaciones en la Isla Iquitos (R. Amazonas) demuestran que también los frailes consumen los choclos (L. Moya, com. pers.) y plátanos. Kurnmer (1973) sostiene que tal adaptación se transmite de generación en generación en la tropa. Otras manadas que habitan los bosques inundables de Nueva York, Palizada y Cocha Sapote A, ingresan a los terrenos cultivados y no muestran interés por los choclos y plátanos expuestos y comidos parcialmente por otros consumidores oportunistas, tal como se mencionó anteriormente. Los terrenos cultivados de Nazareth y Cocha Sapote (A, B, C) tenían menos de dos años de antigüedad; mientras que en Nueva York y Palizada las chacras contaban con menos de un año. Por consiguiente, la adaptación al consumo es gradual y progresiva (Cuadro 3, Fig. 2).
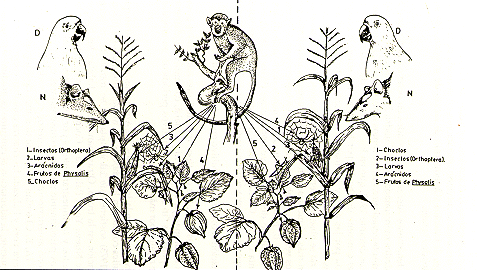
Fig. 3. - Comportamiento alimenticio de
Saimiri spp. ante cultivos dee Zea mays
en terrenos inundables
Las manadas de frailes del tipo gótico de los bosques de Puerto Miguel, donde existen cultivos antiguos, manifiestan cierto grado de aceptación y consumo de choclos y plátanos ; entre tanto, otras manadas de Nueva York, Cocha Urcu, Santa Cruz, Bombonaje, Bellavista, y San Martín, no ingresan (u ocasionalmente atraviesan) a los cultivos de maíz y plátano. Estos terrenos tenían entre uno a dos años (Cuadro l).
5. Escasez de alimentos (o frutos) en las áreas naturales:
De Junio a Noviembre, la escasez de frutos silvestres determina el incremento de la actividad diaria de búsqueda de alimentos de los frailes en el bosque bajo, "purmas"(Denominación que se da a terrenos con vegetación secundaria) y terrenos con cultivos periódicos; los choclos, plátanos y otros frutos cultivados son incluidos en su dieta frugívora (Cuadros 4 y 5) por la inmediata y amplia disponibilidad (Baldwin y Baldwin 1976), convirtiendo a los frailes en competidores naturales o plagas del hombre.
Los ejemplares cautivos procedentes de Palizada, Cocha Sapote (A, B, C), Puerto Miguel, Cocha Urcu (A, B), no consumieron los choclos sino después de dos a cinco días de atrapados; mientras que los procedentes de Sarapanga, Yucuruchi y Huaisi, los consumieron inmediatamente, notándose además en ambos grupos, la inmediata y casi uniforme aceptación de los plátanos. Comparativamente, después de cinco días, todos los grupos cautivos consumieron tanto los choclos como los plátanos, manifestando uniformidad en el consumo de ambos frutos después de 10 días. Esto confirma el rápido acondicionamiento y adaptación del fraile a fuentes de alimentos palatables desconocidos en su área (Kummer 1973).
CONCLUSIONES
El incremento de la actividad agrícola orientada al cultivo de maíz y plátano, durante los últimos tres años, disminuye las áreas de hábitats naturales de los frailes, induciéndolos en la adaptación a nuevas fuentes de alimento.
El maíz, el plátano y otros frutos cultivados aumentan la diversidad alimenticia de los frailes, los que se convierten entonces en plagas agrícolas, compitiendo con los pobladores ribereños. Es necesario efectuar una evaluación de las interrelaciones económicas, del rendimiento de los cultivos diversos y el manejo extensivo de los frailes.
El maíz y el plátano son frutos introducidos en la agricultura ribereña de la amazonía, que requieren del manejo agrícola, mientras que la uvilla, el caimito, la guava, el shimbillo, etc., son frutos nativos o naturalizados que soportan las condiciones semiculturales del bosque en recuperación (o purma) durante algunos años.
Los frailes aprenden gradual y progresivamente a consumir choclos, plátanos y otros frutos exóticos y palatables, inducidos por : a) Otros animales silvestres frugívoros, b) la amplitud (extensión) e intensidad de los cultivos; e) el tiempo transcurrido desde el inicio de la actividad agrícola destinada al cultivo de maíz, plátano, etc.; d) la escasez temporal de frutos silvestres que conforman su dieta natural.
Los frailes son de comportamiento alimenticio insectívoro y frugívoro, dedicándose a esta última actividad principalmente en horas matutinas y vespertinas, en cuyo lapso invaden los terrenos cultivados, motivados por el menor efecto de la insolación y mayor humedad atmosférica.
AGRADECIMIENTO
Expreso mis agradecimientos a los directivos del IVITA y de la Estación de Conservación y Reproducción de Primates No Humanos de Iquitos, a la Organización Panamericana de la Salud, a los funcionarios de la Dirección Regional de Agricultura y Alimentación del ORDELORETO, por todas las facilidades brindadas. A los Drs. Gerardo Lamas, del Museo de Historia Natural "Javier Prado", Lima, y Carlos Málaga, Consultor OPS, por la revisión del manuscrito y sugerencias. Igualmente, a los Srs. Nemesio Collantes (Tahuayo), Nemesio Cahuaza (San Joaquín de Omagua) y Oswaldo Tamani (Nauta), quienes actuaron como guías de campo y atrapadores.
REFERENCIAS
Baldwin, J.D. y J.1. Baldwin. 1976. Primate populations in Chiriqui, Panama. pp. 2031 En R.W. Thorington y P.G. Heltne (Eds.). Neotropical Primates. Field studies and Conservation. National Academy of Sciences, Washington, D.C.
Cain, S.A, G.M. de Oliveira Castro, J. Murca. Pires y N.T. Da-Silva. 1956. Application of some phytosociological techniques to Brazilian Rain Forest. Am. I.Bot. 43:911-941.
Encarnación, F. 1985. Introducción a la flora y vegetación de la Amazonía Peruana: estado actual de los estudios, medio natural y ensayo de una clave de determinación de las formaciones vegetales en la llanura amazónica. Candollea 40: 237252.
Kummer, H. 1973. Primate Societies. Group techniques of ecological adaptation. Aldine Publishing Company, Chicago.
Malleux, J. 1975. Mapa Forestal del Perú (Memoria Explicativa). Universidad Nacional Agraria La Molina, Lima.
Marlier, G. 1979. Limnology of the Congo and Amazon Rivers. pp. 223-238. En Bj. Meggers, E.S. Ayaensu y W.D. Duckworth (Eds.). Tropical Forest Ecosystems in Africa and South America. A Comparative Review. Smithsonian Inst. Press, Washington, D.C.
Meggers,B.J. 1973. Some problems of cultural adaptations in Amazonia with emphasis on the pre-European period. pp. 311-320. En Bj. Meggers, E.S Ayaensu y W.D. Duckworth (Eds.). Tropical Forest Ecosistems in Africa and South America. A Comparative Review. Smithsonian Inst. Press, Washington, D.C.
Oficina Nacional de Evaluación de Recursos Naturales. (ONERN). 1976. Mapa Ecológico del Perú y Guía Explicativa.
Prance, G.T. 1979. Notes on the vegetation of Amazonia. III The terminology of Amazonian types subject to inundation. Brittonia 31(l):26-38.
Rumbaugh, D.M. 1968. The learning and sensory capacities of the squirrel monkey in phylogenetic perspective. pp. 255-317. En L. A. Rosenblum y R.W. Cooper (Eds.). The Squirrel Monkey. Academic Press, New York.
Sioli, H. 1974. Tropical rivers as expressions of their terrestrial environments. pp. 275-288. En F.B. Golley y E. Medina (Eds.). Tropical Ecological Systems Trends in Terrestrial and Aquatic Research. Springer Verlag, New York.
Soini P. y L. Moya. (este volumen). Informe sobre los resultados de los estudios primatológicos en la Zona Norte de la Isla Iquitos.
Thorington, R.W. 1968. Observations of squirrel monkey in a Colombian forest. pp. 6985. En L.A. Rosenblum y R.E. Cooper (Eds.). The Squirrel Monkey. Academic Press, New York.
Wilson, E.O. 1975. Sociobiology. The new synthesis. Harvard
University, Massa
