

![]()
Principal Types and Forms of Sleeping Sites
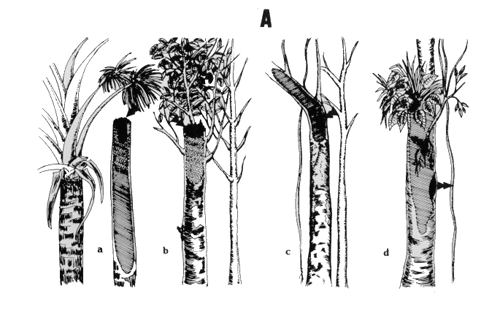
Sleeping sites included 1) holes and concavities in trees, 2) climbers and vines supported by trees, 3) masses of hemiepiphytes and hemiparasties, and 4) thickets and dense branches of shrubs and bamboos (Fig. 2). These four primary types of sleeping sites comprised eleven different or variant forms.
Type A. Sleeping sites are in holes of tree trunks. Access is by the thin trunks and branches of neighboring trees, climbers, or vines. This type of sleeping site affords optimal conditions of shelter and security from predators.
Form Aa. This form is the apical position in the dry trunk of a palm (generally Scheelea spp., Guilielma gasipaes, or Mauritia flexuosa), whereby entrance is gained by the trunk or.some proximal leaf. The superposition of branches and leaves of adjacent trees prevents the direct entry of rainwater into the hole (Fig. 2Aa).
Form Ab. This form is the apical position in the trunks of dry or senescent trees that can have "reiterations" [sensu Halle et al, 1978]. The entry of rainwater into the hole is prevented by the superposition of the branches of taller neighboringtrees (Fig. 2Ab).
Form Ac. This form is the subaxial position on senescent or dry trunks where the entrance corresponds to the knot of a fallen branch (Fig. 2Ac).
Form Ad. This form is the subapical position with one or more entrances in dry or senescent trunks. Woody epiphytes whose roots reach into the cavity and form steps or shelves are used by Aotus for resting or daytime sleep (Fig. 2Ad).
Form Ae. This form is the lateral position over thick, dry, or senescent branches of trees Macrolobium sp., Eschweilera sp., and Calycophyllum spruceanum (Fig. 2 Ae).
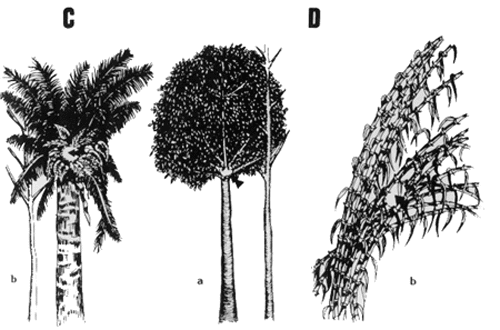
Type B. These complex sleeping sites are in the concavities of branches sup porting entanglements of climbers and vines, which serve as access routes and provide protection and shelter. Branching patterns are principally of the Koriba, Prevost, or Nozeran models [Halle et al, 1978].
Form Ba. This is an axial concavity formed by more than four branches (Fig. 2 Ba).
Form Bb. This is a subaxial concavity formed by the loss or fall of a branch (Fig. 2Bb).
Type C. These complex sleeping sites are composed principally of epiphytes, hemiepiphytes, and hemiparasites (Bromeliaceae, Araceae, Loranthaceae, Guttiferae, Moraceae, Gesneriaceae, ferns, and others), almost always in parabiosis with ants. Access is by vines or by the branches of neighboring trees.
Form Ca. This form is the subaxial position over a slight concavity protected by diverse epiphytes and hemiepiphytes. The concavity is produced by the fall of a branch in the Nozeran model [Halle et al, 1978] (Fig. 2Ca).
Form Cb. This form is the subapical position without concavity. The sleeping site is at the base of the diverse epiphytes and hemiepiphytes that grow on a palm of the intermediate model of Holttum and Corner [Halle et al, 1978] (Fig. 2Cb).
Form D. These simple sleeping sites are positioned among thickets and dense foliage with multiple or defined access.
Form Da. This form is the axial position with protection provided by the foliage of branches of trees principally in the models of Aubreville, Mangenot, or Massart [Halle et al, 1978] (Fig. 2Da).
Form Db. This form is the irregular position with protection from foliage and thickets of dense shrubs or trees with poliaxial branching, as occurs in bamboos of the McClure model or Myristicaceae of the Aubreville model [Halle et al, 1978] (Fig. 2Db).
The form Ac, characterized by the presence of epiphytes and woody hemiepiphytes (Ficus sp., Coussapoa sp., Clusia sp., Souroubea sp., and others) with root systems rooted in the holes, can be confused with form Bb when they are covered by herbaceous or semiwoody climbers (Bignoniaceae, Vitaceae, Cucurbitaceae, Menispermaceae, Convolvulaceae, and others) and some succulent and herbaceous epiphytes.
In dry or senescent trees, the orientation of the cavity is generally descending with respect to the entrance or opening. However, in the forms Aa and Ad, it can occur with an ascending orientation. In forms Ba and Bb, protection from rain is complemented by the foliage, the decomposition and detachment of cork, and the disintegration of arthropod and other remains, which are sometimes the substrate of certain epiphytes (Araceae, Bromeliaceae, Cyclanthaceae, Gesneriaceae, etc).
In form Ca, the foliar scars and the spathes of palms such as Scheelea cephalotes, S. bassleriana, and others called "shapaja" constitute the base of the sleeping site, whose form is then complemented with Philodendron sp., Ficus sp., Polypodium sp., and Nephrolepis sp. among the epiphytes.
The form Da is frequent among small trees of Rheedia sp., Ocotea sp., Iryanthera sp., Cordia sp., Sapium sp., and others, while the form Db is found over thickets of Guadua superba in Loreto Department and over Guadua sp. between the Manuripe and Acre Rivers in Madre de Dios Department.
Description of Sleeping Sites
Tree holes were the most common sleeping sites of Aotus nancymai and A. vociferans. These cannot, in any way, be properly considered built nests. Other nest types, which lacked walls, were also unmodified. Tree holes were characterized by one to three orifices or access routes into the internal cavity. We have rarely found holes with four or five orifices and have found them only in trees of Calycophyllum spruceanum, Campsiandra laurifolia, and Eschweilera sp. However, only one or two of these routes were used by Aotus for routine entries and exits; the others were used only circumstantially. Access holes were generally almost circular, with diameters varying between 14 and 30 cm.
The internal cavity was generally cylindrical in form, having the same or a somewhat larger diameter than the orifice. The depth of the cavity varied between 0.6 and 13.4 in. Some cavities contained roots of hemiepiphytes and hemiparasites that formed steps on ladders for Aotus . Generally, the cavity was clean and more or less dry. Organic remains, if they existed, were found at the bottom of the cavity in the process of decomposition, These remains served as substrate for fly larvae and other insects. The walls of the internal cavity and orifice showed wear and polish as a result of the constant traffic of individuals. These walls were impregnated with odoriferous substances secreted by the subcaudal and pectoral glands of Aotus [Aquino & Encarnación, 1986].
Trees Containing Holes
Nest holes were found in 67 old and senescent trees comprising 13 species. Among these, we found 40.2% of the sleeping sites in trunks and branches of Eschweilera sp., 12.0% in Macrolobium acaciaefolium, 10.3% in Campsiandra laurifolia, and 9.0% in Calycophyllum spruceanum (Table 1). These tree species characteristically show disintegration of the heartwood even though the trunk, supported only by sapwood, remains erect and partially vegetative.
Distribution of Sleeping Sites With Respect to Forest Strata
The ages of individual trees and their densities and crown shapes were factors that determined the location and selection of sleeping sites. In interpreting the location of these sites, the following forest strata were identified: understory (7.110.0 m), lower story (10.119.0 m), middle story (19.125.0 m), upper story (25.131.0 m), and emergent trees (31.137.0 m).
In the forest of the Tahuayo River, 21.4% of the sleeping sites of Aotus nancymai were in the shrub stratum of the understory, 64.3% were in the lower story, and 14.3% were in the middle story. No sleeping sites were found in the upper story or in emergent trees.
In contrast, in the forest of the Nanay and Napo Rivers, 11.4% of the sleeping sites of A. vociferans were in the understory, 54.3% were in the lower story, 20.0% were in the middle story, 8.6% were in the upper story, and 5.7% were in emergent trees (Table II; Fig. 3).
Each forest stratum had different forms of sleeping sites. The Aa, Da, and Db forms were found in the understory; the Ab, Ac, Ba, and Bb forms were found in the lower story; and the Ad, Ae, Ca, and Cb forms were frequently seen in the highest strata (Figs. 2 and 3).
Competition Over and Sharing of the Sleeping Sites With Other Mammals
In Aotus nancymai and A. vociferans, we observed competition over and sharing of the type A sleeping sites with other mammals of similar nesting habits. The size and depth of the holes were important in these interactions. We observed one case of competition and six cases of cooccupation in several circumstances and localities. Moreover, bats often roosted in these cavities.

Fig. 2. Types and forms of sleeping sites. A. Sleeping sites in holes. As. Apical position in palm trunk. Ab. Apical position in dicotyledenons tree trunk. Ac. Subaxial position. Ad. Subapical position. Ae. Lateral position. B. Complex sleeping sites in concavities with climbers and vines. Ba. Axial concavity. Bb. Subaxial concavity. C. Complex sleeping sites with hemiepiphytes and epiphytes. Ca. Subaxial position. Cb. Subapical position. D. Simple sleeping sites in thickets and dense foliage. Da. Axial position. Db. Irregular position.
TABLE 1. Trees With Aotus Nest Holes Observed
in Amazon Lowlands of Peru.
| Species |
Vernacularnames
|
Frequency
|
Percentage
|
| Guilielma gasipaes |
Pijuayo
|
3
|
4.5
|
| Calycophyllum spruceanum |
Capirona
|
6
|
9
|
| Campsiandra laurifolia |
Huacapurana
|
7
|
10.3
|
| Clarisia biflora |
Capinuri
|
2
|
3
|
| Coccoloba sp. |
Vino Huayo
|
1
|
1.5
|
| Couroupita subsessilis |
Ayahuma
|
2
|
3
|
| Eschweilera sp. |
Machimango
|
27
|
40.2
|
| Ficus sp. |
Renaco
|
2
|
3
|
| Hura crepitans |
Catahua
|
2
|
3
|
| Iriartea sp. |
Huacrapona
|
3
|
4.5
|
| Macrolobium acaciaefolium |
Pashaco negro
|
8
|
12
|
| Manilkara sp. |
Quinilla
|
1
|
1.5
|
| Mauritia flexuosa |
Aguaje
|
3
|
4.5
|
| Totals |
|
67
|
100
|
In the locality of Mishana on the Nanay River, for a family group of Aotus vociferans with three individuals we found four sleeping sites, one of which was used for 3 consecutive days. At 0525 hours on the 4th day, an adult Potos flavus entered the hole; 17 minutes later the Aotus group arrived but, hearing growls from within the cavity, did not enter and moved to another sleeping tree located 30 in away. After that incident, the Aotus group used two other sleeping trees for their diurnal rest, without returning to the first, even after it was abandoned by the P. flavus, which used it during 4 consecutive days.
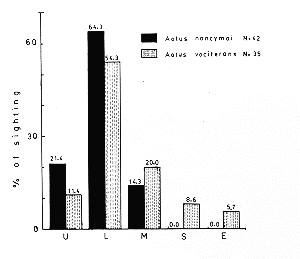 |
Fig. 3. Percentage distribution of Aotus nancymai and A. vociferans sleeping sites by forest strata. U, understory; L, lower story; M, middle story, S, upper story; E, emergent trees. |
Of the six cases of cohabitating, two were with Potos flavus, one was with Bassaricyon gabbi, and three were with Coendu bicolor. Aotus always occupied that part of the cavity nearest the entrance, while the interiormost part was occupied by the other species. All these cooccupants entered the sleeping sites between 15 to 20 minutes before Aotus and left between 20 and 25 minutes after Aotus .
These results indicate that competition occurred when the sleeping site was small and shallow and had only one entrance; cooccupation occurred when the sleeping site was large and deep and had two or more entrances, and where root mats of hemiepiphytes and hemiparasites formed lattice works used as substrata by the occupants.
TABLE II. Frequency Distribution of the Sleeping Sites by Forest Strata A. nancymai
| .Stratum | .
Height (M)
|
A. nancymai
|
A. vociferans
|
||
|
Frequency
|
Percentage
|
Frequency
|
Percentage
|
||
| Understory |
7.1-10.0
|
7
|
21.4
|
4
|
11.4
|
| Lower story |
10.1-19.0
|
27
|
64.3
|
19
|
54.3
|
| Middle story |
19.1-25.0
|
6
|
14.3
|
7
|
20
|
| Upper story |
25.1-31.0
|
0
|
0
|
3
|
8.6
|
| Emergent trees |
31.1-37.0
|
0
|
0
|
2
|
5.7
|
| Totals |
|
42
|
100
|
35
|
100
|
